Чем хорошо авторство в биореакторе, так это тем, что в процессе подготовки заметки, можно узнать массу нового. И иногда это новое, настолько отличается от имеющихся знаний, что приходится отказываться от старого.
Вот, например, нам задали вопрос, как получаются сиамские близнецы – казалось бы чего проще, ведь уже много десятилетий известно, что… А что, собственно, известно много десятилетий? Беглое знакомство с вопросом по данным выдачи в поисковике никакой ценной информации не дал. А в литературных источниках из книги в книгу перекочёвывает одна и та же гипотеза, согласно которой образование сиамских близнецов связано с «нарушением миграции бластомеров на ранних этапах эмбриогенеза». В принципе, тут можно было бы поставить жирную точку и расходиться. Но всё не так просто – что за бластомеры? Почему произошло нарушение миграции? И, почему, чёрт возьми, эта миграция вообще должна происходить?
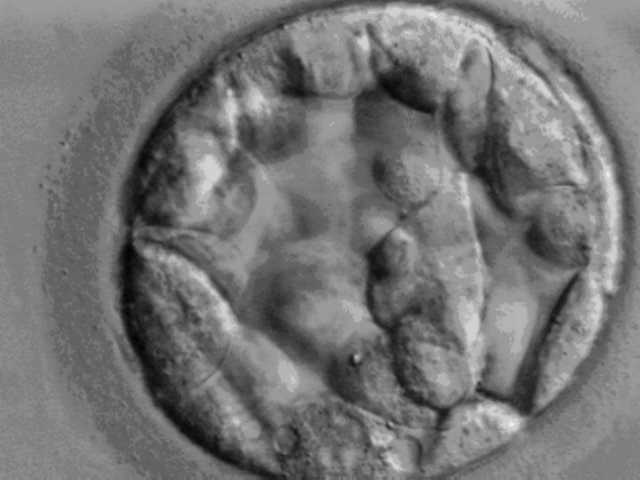
Витрины музеев и кафедр патологической анатомии и судебной медицины буквально ломятся от разного рода «уродцев», а понимания, почему это происходит не прибавляется – давайте попробуем вместе разобраться в этом вопросе.
Для начала стоит понять, как в норме происходит образование близнецов. Тут стоит договориться о некоторых дефинициях – близнецы бывают двуяйцевым и однояйцевыми. В первом случае, всё просто – два сперматозоида оплодотворили две яйцеклетки (иногда случается так, что во время овуляции созревает не одна, а две яйцеклетки). И получилось два разных организма. Во втором случае всё интересней – два организма получаются в результате того, что отдельные клетки – бластомеры, уже оплодотворённой яйцеклетки – зиготы, по тем или иным причинам потеряли связь друг с другом и превратились в два независимых, но почти идентичных, организма.
Первые опыты по разделению отдельных бластомеров и наблюдениями, что из этого получится, эмбриологи начали проводить ещё в конце XIX – начале ХХ века. Опыты, главным образом, осуществлялись на морских ежах, амфибиях (тритонах и лягушках) и, гораздо реже – на высших позвоночных. Выяснилось, что вероятность того, что из разделённых бластомеров получится новый организм, обратно пропорциональна степени организации животного. То есть, морулу («комок» клеток, получившихся в результате дробления зиготы) морского ежа, можно разделять на отдельные клетки довольно долго, и из каждой клетки получится личинка, которая станет взрослым организмом, лягушки или тритона – гораздо меньше, а вот у высших позвоночных, возможности появления зародыша исчерпываются стадией всего 8 клеток (3-е дробление зиготы). Если разделить бластомеры на 4-ом дроблении, все они погибнут. Дополнительной прелести ситуации придает и то, что яйцеклетка млекопитающих покрыта т.н. блестящей зоной или zona pellucida – тонкой «плёнкой», состоящей из гликопротеидов, которая препятствует проникновению более чем одного сперматозоида (полиспермии), а после оплодотворения служит «каркасом», который удерживает бластомеры на ранних этапах развития (Рис. 1).
Тут-то мы и сталкиваемся с первым парадоксом – бластомеры млекопитающих не способны образовывать прочные межклеточные взаимодействия до 4-ого деления, и, если блестящая оболочка разрушается раньше – велика вероятность образования однояйцевых близнецов. Кстати, у некоторых позвоночных, а, конкретнее, у броненосцев, механизм разрушения блестящей оболочки и расхождения бластомеров обусловлен эволюционно - у них почти во время каждой беременности, получается 8 однояйцевых близнецов.
Но как тогда могут образовываться сиамские близнецы? Получается, что бластомеры успели образовать какие-никакие межклеточные контакты, но всё ещё способны «жить независимо». Выходит, какое-то дробление 3,5 (платформа 9 и 3\4, ага). Тем не менее, это не смущало эмбриологов, и длительное время считалось, что именно это – нарушение расхождения независимых бластомеров приводило к появлению сиамских близнецов. Тем более, что на другом излюбленном экспериментальном эмбриологическом объекте – яйце – всё прекрасно складывалось. Правда у яйца нет блестящей зоны… А вот двухголовые змеи и четвероногие
куры есть.
Но объём наблюдений за сиамскими близнецами накапливался, и он потребовал создать какую-либо классификацию, которая упорядочит имеющуюся информацию.
И такая классификация была создана. В первую очередь, всех сиамских близнецов разделили на симметричных и ассиметричных. Симметричные близнецы представляют собой два равноценных организма, сросшихся между собой в том или ином месте. Для того, чтобы и тут навести порядок, было принято обозначать близнецов по месту сращения, добавляя корень пагус- (от греческого слова – «соединять»). Так получились краниопаги – близнецы, сросшиеся головами, торакопаги – сросщиеся в области груди и пигопаги – сросшиеся в области таза (Рис. 2).
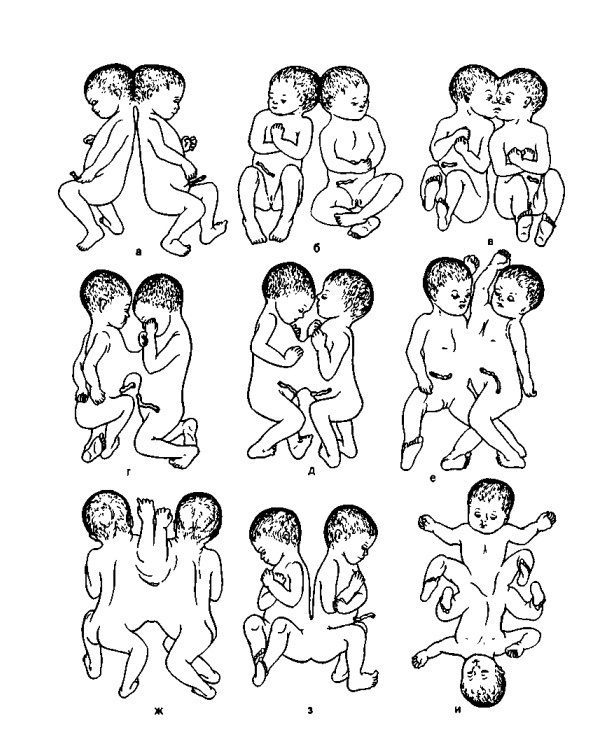
Таких близнецов, собственно, и называют сиамскими – в честь Энга и Чанга Банкеров – широко известных близнецов, которых демонстрировал Финеас Барнум в своём знаменитом цирке (Рис. 3). Справедливости ради стоит заметить, что это не первые сиамские близнецы в истории человечества – так, хроники сообщают, что при дворе шотландского короля Иакова IV, жившего в XVI веке, был человек, у которого над общим тазом и ногами возвышалось две грудные клетки и две головы. Похожее свидетельство можно найти и в истории крестовых походов, где описан лучник из войска Саладина, обладавший двумя головами. Тем не менее, первыми «задокументированными» близнецами стали Энг и Чанг из Сиама, которые и дали название для всего явления.
В противоположность симметричным, существуют ассиметричные близнецы. В этом случае только один индивид является полноценно сформированным, а другой (второй, третий… максимум известно 4 - рис. 4) представляется собой недоразвитый придаток, своего рода «паразита». Их так и называют – полноценного – «аутозитом», а менее полноценного – «паразитом». Первым задокументированным случаем такого рода стало рождение в 1617 году в Генуе графа Лаццаро Колоредо, на груди которого располагался рудиметарный близнец (Рис. 5). «Паразит» достиг 90 см длины, имел чётко оформленную голову, две трёхпалые руки и одну левую ногу. Иногда он даже двигал руками и шевелил губами. При этом рудиментарный близнец дышал самостоятельно и у него выслушивались собственные тоны сердца.
Похожий случай описал великий немецкий патолог Рудольф Вирхов на случае индуса Лалоо, у которого из груди рос «паразит», лишённый головы, который, по сути, представлял собой только нижнюю половину тела (Рис. 6).
Однако не стоит думать, что такие ассиметричные близнецы обязательно должны себя проявлять. В ряде случаев, близнец-паразит полностью расположен в полостях тела, и его обнаруживают только при инструментальном исследовании. Такой тип ассиметричных близнецов, иногда называют матрёшкой. В мировой литературе с 1808 по 1978 описано 28 подобных случаев.
Именно асимметричные близнецы долгое время ставили эмбриологов в тупик – если бластомеры разошлись раньше, чем разрушилась блестящая оболочка, то почему в одних случаях образуются симметричные близнецы, а в других – нет.
Для объяснения этого феномена была предложена другая гипотеза, согласно которой ассиметричные близнецы образуются в результате поглощения, ассимиляции, одного эмбриона (более мелкого) – вторым – более крупным. В свою очередь, нарушение развития эмбрионов объясняли патологией трофических механизмов во время беременности.
Две эти гипотезы – нарушение расхождения бластомеров до разрушения блестящей оболочки и ассимиляции эмбрионов, просуществовали почти до конца ХХ века. Собственно, в таком виде их преподавали в медицинских университетах ещё два десятка лет назад.
И изменить взгляд на природу сросшихся близнецов помогло одно открытие и одно наблюдение. Первое – открытие группы гомеозисных или НОХ-генов. Что же это за необычные гены?
Гены, и в правду, необыкновенные. Это высококонсервативные гены. Их (с некоторыми изменениями, естественно), но в целом очень похожих, можно обнаружить среди ВСЕХ многоклеточных организмов (у растений, они, правда, довольно сильно отличаются, но суть несут ту же).
Белки, которые кодируют эти гены, определяют процессы роста и дифференцировки в организме. Эти гены,
впрочем, как и все остальные, расположены линейно, но «включаются» они по очереди – т.е. продукты первого в цепочке гена, запускают не только процессы клеточной дифференцировки, но и следующий ген. По сути, именно работа этих генов определяет, где у эмбриона будет голова, где передние конечности, где – задние, а где хвост. По одной из гипотез, именно мутации в этих генах приводят к макроэволюционным изменениям. Например, увеличение числа определённых генов из этой группы привело к тому, что у змей увеличилось число позвонков, а у насекомых из одинаковых конечностей сформировались разные анатомические структуры. Так что, в некотором роде, гомеозисные гены – это общий «чертёж» организма.
И тут мы переходим к наблюдению. В настоящий момент известно более 2000 случаев удвоения голов у змей (в 227 разных видах) и около 100 – у черепах (в 30 разных видах). Но самое главное, что в некоторых из этих наблюдений, число позвонков и рёбер у разных «половин» могло отличаться! А это значит, что тут не обошлось без гомеозисных генов.
Но потребовалось еще несколько лет, прежде чем исследователи смогли расшифровать часть из сигнальных каскадов, которые запускают эти гены.
В настоящее время считается, что среди гомеозисных генов есть три основные группы – те, которые определяют формирование организма по «переднезадней оси», по оси «право-лево» и по оси «спина-живот».
И сейчас немного эмбриологии. Из курса общей биологии весьма средней школы мы помним, что одним из этапов эмбионального развития является гаструляция. В учебниках, как правило, её описывают как процесс впячивания внутрь себя пузырька – бластулы, с образованием двуслойного зародыша, первичной полости тела – бластоцеля, и полости первичной кишки – которая сообщается с внешней средой посредством первичного рта – гастропора. Эта картинка соответствует действительности, но только для ланцетника. У высших позвоночных и, в частности, у млекопитающих, все происходит несколько иначе.
У млекопитающих гаструляция происходит в два этапа, первый - деламинация – расслоение клеток на эпибласт и гипобласт. Второй – миграция – клетки эпибласта начинают смещаться, образуя первичную полоску и гензеновский узелок. Клетки, остающиеся на поверхности, сформируют эктодерму, а мигрирующие вглубь – мезо- и эндодерму. Но нас интересует именно гензеновский узелок – с точки зрения эволюции – это гомолог дорзальной – спинной – губы бластопора. Из него под действием продуктов гомеиозисных генов образуется нервный тяж (Рис. 8).
Всем этим «ансамблем» управляют гомеозисные гены. В случае, если мутация произошла в одном из них, возможно появление, например, второго гензеновского узелка, вокруг которого сформируется вторая хорда, вокруг второй хорды – вторые осевые органы, и т.д. В зависимости от того, в каком из НОХ-генов произошла мутация – на таком уровне и будут срощены близнецы.
Но это ещё не всё – близнецы могут быть срощены разными поверхностями тела, например спина к спине, или наоборот – грудь к груди. Кроме этого, внутренние органы сросшихся близнецов расположены зеркально относительно друг к друга, т.е. у одного близнеца сердце будет слева и левая дуга аорты, а у второго – справа и правая дуга аорты. И это обусловлено результатом работы других гомеозисных генов (Рис. 9).
В углублении гензеновского узелка, расположены специфические клетки, поверхность которых покрыта ресничками. По одной из гипотез, под действием тока жидкости эти реснички наклоняются, в результате чего выделяются специфические вещества – сигнальные молекулы, которые определяют формирование право-левой оси организма. Этим процессом управляет ген Nodal. В случае, если реснички отсутствуют или «работают» неправильно, происходит нарушение деятельности гена Nodal, что приводит к нарушению формирования право-левой симметрии. У одного эмбриона это приводит к формированию разного рода аномалий, вроде инверсии расположения внутренних органов разной степени тяжести, а у сиамских близнецов – к зеркальному расположению внутренних органов (Рис. 10).
Для многих рептилий (ящерицы, змеи, крокодилы и черепахи) показано,
что причиной нарушения работы сигнальных путей, определяющих формирование гензеновского пузырька может стать воздействие высокой температуры во время инкубации яиц. Для млекопитающих такой причиной может стать воздействие ионизирующей радиации и некоторых химических соединений. Тем не менее, окончательная точка в вопросах причин и механизмов появления сросшихся близнецов не поставлена.
Кто знает, как изменятся знания человека об эмбриогенезе ещё через два десятка лет.
Список использованной литературы:
1. Акушерство: Учебник/ Г.М. Савельева. В.И. Кулаков, А.Н. Стрижаков и др.; Под ред. Г.М. Савельевой. – М.: Медицина, 2000
2. Балахонов А.В. Ошибки развития. – Л.: Изд-во Ленинград. Ун-та, 1990
3. Барреси М. Дж. Ф. Биология развития / М.Дж. Ф. Барреси, С.Ф. Гилберт ; пер. с англ. под ред. др-а биол. Наук А.В. Васильева. – М.: Лаборатория знаний, 2022
4. Ермаков А.С. Правая, левая, где сторона? /Ермаков А.С. - //Химия и жизнь. – 2014. - №8
5. Журавлёв А.Ю. Как живые: Двуногие змеи, акулы зомби и другие исчезнувшие животные Э Андрей Журавлёв. – М.: Альпина нон-фикшн, 2024
6. Касимова, Р. М. Установление лево-правой висцеральной асимметрии в эмбриогенезе человека / Р. М. Касимова, А. Д. Тимошок. — // Молодой ученый. — 2019. — № 48 (286).
7. Пэттэн Б.М. Эмбриология человека / Пер. с англ. О.Е. Вязова и Б.В. Конюхова. Под ред. Проф. Г.А. Шмидта – М.: МедГиз, 1959
Автор: Артемий Липилин