Все аэробные организмы подвержены определенному уровню физиологического окислительного стресса в результате митохондриального дыхания.
Образующиеся промежуточные продукты, такие как супероксид (O2-•) и перекись водорода (H2O2), могут привести к образованию токсичных радикалов кислорода, которые могут вызвать перекисное окисление липидов и повреждение клеток. Чтобы предотвратить это, эндогенно образующуюся перекись водорода восстанавливают с помощью GSH в присутствии селен-зависимой GSH-пероксидазы (рис. 3).
При этом GSH окисляется до GSSG, который, в свою очередь, восстанавливается обратно до GSH редуктазой GSSG за счет НАДФН, образуя окислительно-восстановительный цикл. Органические пероксиды могут быть восстановлены с помощью GSH-пероксидазы и GSH-S-трансферазы.
Перекись водорода также может быть восстановлена каталазой, которая присутствует только в пероксисоме. В митохондриях GSH особенно важен, поскольку здесь нет каталазы. Действительно, митохондриальный GSH имеет решающее значение для защиты как от физиологически, так и от патологически вызванного окислительного стресса (Fernandez-Checa et al., 1997; Garcia-Ruiz and Fernandez-Checa, 2006).

GSH как хранилище цистеина и цикл γ-глутамила
Хранение цистеина является одной из наиболее важных функций GSH, поскольку цистеин чрезвычайно нестабилен внеклеточно и быстро самоокисляется до цистина в процессе, который приводит к образованию потенциально токсичных свободных радикалов кислорода (Meister, 1988).
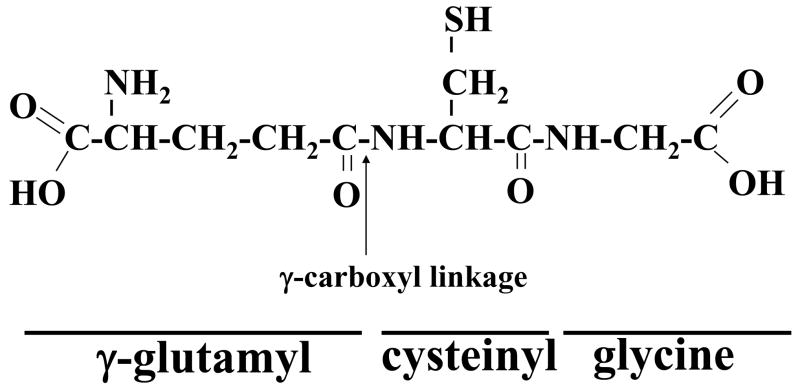
Мейстер впервые описал цикл γ-глутамила в начале 1970-х годов, который позволяет GSH служить постоянным источником цистеина (рис. 4). В этом цикле GSH высвобождается из клетки с помощью переносчика(ов), а эктофермент GGT затем переносит γ-глутамиловую часть GSH на аминокислоту (лучшим акцептором является цистин), образуя γ-глутамиловую аминокислоту. и цистеинилглицин. Затем γ-глутамиламинокислота может быть перенесена обратно в клетку для завершения цикла. Попав внутрь клетки, γ-глутамиламинокислота может подвергаться дальнейшему метаболизму с высвобождением аминокислоты и 5-оксопролина, которые могут быть преобразованы в глутамат и использованы для синтеза GSH.
Цистеинилглицин расщепляется дипептидазой с образованием цистеина и глицина. Большинство клеток легко поглощают цистеин. Попав внутрь клетки, большая часть цистеина включается в GSH, некоторая часть включается в белок, в зависимости от потребностей клетки, а часть расщепляется до сульфата и таурина (Meister, 1988). Для большинства клеток цикл γ-глутамила позволяет использовать GSH в качестве постоянного источника цистеина.
GSH как регулятор роста и гибели
В таких типах клеток, как лимфоциты и фибробласты, повышенный уровень GSH связан с ранним пролиферативным ответом и необходим для перехода клетки в S-фазу (Shaw and Chou, 1986; Messina and Lawrence, 1989; Hamilos et al., 1989). ; Ивата и др., 1994; Пут и др., 1995). Мы также сообщили о повышении уровня GSH во время пролиферации гепатоцитов крысы, например, при посеве первичных культур гепатоцитов крысы при низкой плотности, что стимулирует гепатоциты переходить от фазы G0 к фазе G1 клеточного цикла (Lu and Ge., 1992), а также после 2/ 3 частичная гепатэктомия (ПГ) (Huang et al., 1998). Увеличение уровня GSH в печени после ЛГ происходило за счет усиления биосинтеза и до начала синтеза ДНК (Huang et al., 1998). Если это увеличение GSH блокировалось, регенерация печени нарушалась (Huang et al., 2000a). Более того, статус GSH напрямую коррелировал с ростом раковых клеток печени (Huang et al., 2000a). Карретеро и его коллеги также показали, что повышенное содержание GSH способствует росту метастатических клеток меланомы в печени (Carretero et al., 1999). Молекулярный механизм того, как GSH модулирует пролиферацию клеток, остается в значительной степени спекулятивным. Известно, что GSH модулирует синтез ДНК путем поддержания восстановленного уровня глутаредоксина или тиоредоксина, которые необходимы для активности рибонуклеотидредуктазы, фермента, ограничивающего скорость синтеза ДНК (Holmgren, 1981).
Альтернативно, увеличение содержания клеточного GSH может изменить тиол-редокс-статус клетки, что, в свою очередь, может повлиять на экспрессию или активность факторов, важных для развития клеточного цикла.
GSH также участвует в модуляции гибели клеток.
Апоптоз, один из крайностей непрерывного процесса гибели клеток, характеризуется такими морфологическими особенностями, как конденсация и фрагментация хроматина, а также межнуклеосомное расщепление ДНК; другой крайностью является некроз, который характеризуется разрывом или фрагментацией плазматической мембраны и истощением АТФ (Гарсия-Руис и Фернандес-Чека, 2007). Обе формы гибели клеток часто сосуществуют и имеют общие пути, например, с участием митохондрий (Lemasters, 2005). GSH модулирует гибель клеток в обеих крайностях. GSH регулирует окислительно-восстановительное состояние специфических тиоловых остатков белков (таких как NFκB, стресс-киназы и каспазы), участвующих в гибели клеток (Garcia-Ruiz и Fernández-Checa, 2007).
Истощение GSH происходит во время апоптоза во многих различных типах клеток, вторично по отношению к увеличению количества активных форм кислорода (АФК), усилению оттока GSH и снижению активности GCL (см. также Посттрансляционная регуляция GCLC) (Hall, 1999; Franklin et al., 2003). ).
В клетках U937 и HepG2 блокирование оттока GSH предотвращало апоптоз, индуцированный пуромицином (Ghibelli et al, 1998), что позволяет предположить, что истощение GSH играет важную роль в инициации апоптоза в некоторых типах клеток. Однако глубокое истощение GSH превращает гибель клеток, вызванную различными агентами, из апоптоза в некроз (Hall, 1999), что позволяет предположить, что очень высокие уровни АФК могут подавлять апоптотический механизм.
Действительно, GSH может влиять на путь гибели клеток, модулируя уровень митохондриальных АФК, как было продемонстрировано в случае фактора некроза опухоли α (TNFα) (Garcia-Ruiz и Fernández-Checa, 2007).
Истощение митохондриального GSH приводит к повышению уровня АФК и активных форм азота, митохондриальной дисфункции и истощению АТФ. Это может превратить апоптозную гибель клеток в некротическую (Garcia-Ruiz and Fernández-Checa, 2007).