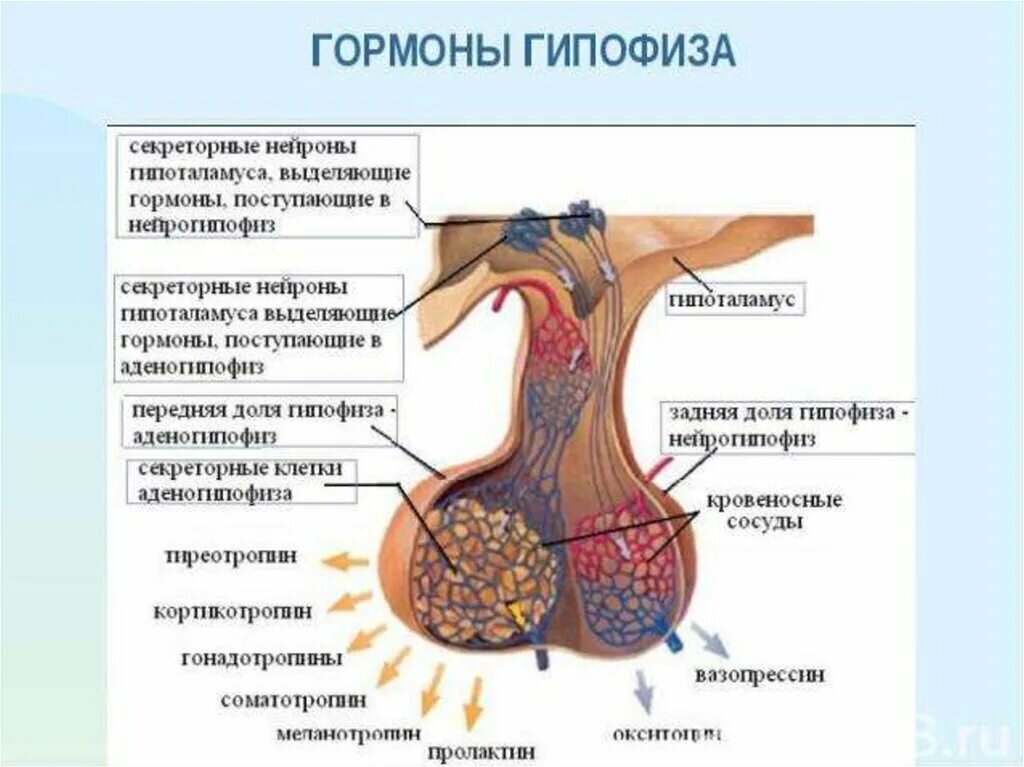
Гипофиз расположен в турецком седле основной кости черепа. С помощью ножки он соединен с гипоталамусом.
Морфологически железа неоднородна. В ней отчетливо выделяется передняя, средняя и задняя доли.
Передняя доля (аденогипофиз) состоит из железистой ткани, вырабатывающей гормоны для регуляции функций других эндокринных желез.
В средней доле секретируется гормон, регулирующий пигментный обЗадняя доля (нейрогипофиз) представлена густой сетью безмякотных нервных волокон и модифицированными клетками нейроглий (питуициты), участвующими в образовании гормонов.
Гормоны передней доли гипофиза (аденогипофиза). Передняя доля ги-
пофиза регулирует главным образом рост и развитие организма, обмен веществ и размножение.
Из передней доли гипофиза выделен ряд гормонов. Некоторые из них получены в химически чистом виде. Главнейшие из этих гормонов следующие:
гормон роста – соматотропный гормон – выделен в кристаллическом виде. Обладает широким диапазоном действия. Он обладает видовой специфичностью, т.е. действует на тот вид животных, у которых вырабатывается.
Например, СТГ животных не действует на человека и наоборот. Этот гормон
стимулирует рост костей и мягких тканей. Считается, что он оказывает главное влияние на костную и хрящевую ткани, участвует в регуляции обмена Са и Р, усиливает синтез рибонуклеиновых кислот и белка. Соматотропный гормон действует и на углеводный обмен. Он усиливает функцию поджелудочной железы, которая выделяет глюкагон в большом количестве, следова-
тельно, СТГ повышает концентрацию сахара в крови. Действует гормон и на жировой обмен – повышает использование и расход жира из жировых депо, с поддерживанием баланса электролитов в организме.
Недостаточная секреция СТГ у животных приводит к карликовости (малый рост и развитие). Повышенная секреция СТГ у молодых животных приводит к гигантизму – усиленному росту, связанному с интенсивным и пропорциональным развитием трубчатых костей.
Если же повышенная секреция гормона (СТГ) происходит у взрослых животных или же вводить взрослым животным избыточное количество СТГ, то наблюдается непропорциональное развитие всех частей тела и органов, особенно отдельных костей лица – резко увеличивается развитие нижней челюсти. Это заболевание носит название акромегалии. Неравномерное разрастание внутренних органов – спланхномегалия. СТГ обеспечивает проявление инстинкта, связанного с заботой о потомстве и регулирует процессы молоко-
образования у самок сельскохозяйственных животных.
Пролактин стимулирует образование молока, действуя на ферментные системы железистых клеток альвеол вымени и увеличивает секрецию прогестерона желтыми телами яичников. Кроме того, пролактин является стимулятором разнообразных процессов, связанных с воспроизведением и воспитанием потомства у голубей. В период вскармливания птенцов в клетках зоба вырабатывается секрет, который голуби отрыгивают в рот птенцам.
Гонадотропные гормоны передней долей гипофиза, влияют на развитие и функцию половых желез, а также на весь организм. Они образуются в базофильных клетках передней доли гипофиза и по своей химической природе представляют глюкопротеиды. Гипофиз как источник гонадотропных гормонов служит главным эндокринным органом, управляющим функциями, связанными с размножением.
Гипофиз молодого млекопитающего может развиваться либо по мужскому, либо по женскому типу в зависимости от того, какая половая железа имеется в организме. У половозрелых млекопитающих гипофиз обнаруживает половую специфичность, т.е. существует различие между женским и мужским гипофизом. Это различие связано с функциональным циклом: если пересадить взрослой кастрированной самке яичники (но не семенники), то тормозится секреция ФСГ гипофизом, а у кастрированного самца секреция ФСГ подавляется только при пересадке семенников.
Передняя доля гипофиза выделяет три гонадотропных гормона: фолликулостимулирующий (ФСГ) гликопротеид с молекулярным весом 30000 –стимулирует у самок рост яичников, созревание фолликулов и образование
половых гормонов – эстрогенов. У самцов этот гормон стимулирует сперматогенез в семенниках до образования спермацитов 1-го порядка.
Другой гормон –лютеостимулирующий (ЛГ) (гликопротеид) с молекулярным весом 32000 способствует овуляции, т.е. выходу яйцеклеток из фолликулов, развитию желтого тела и образованию в нем гормона прогестерона,
а у самцов – тестостерона.
Третий гормон –лютеотропный (ЛТГ) – способствует сохранению жёлтого тела в период беременности и секреции прогестерона, у птиц стимулирует инстинкт насиживания, обеспечивает развитие молочных желез.
Экстракты из передней доли гипофиза у неполовозрелых животных вызывают преждевременное половое созревание. На этой основе разработаны практические мероприятия в области рыбоводства: инъекции суспензий гипофиза самцам и самкам осетровых рыб ускоряют созревание и повышают количество зрелых сперматозоидов, а также способных к оплодотворению и к дальнейшему развитию икринок.
Таким образом, гонадотропные гормоны, оказывают влияние на развитие и функции половых желез.
Меланоцитостимулирующий гормон (МСГ) – полипептид вырабатывается промежуточной (средней) долей аденогипофиза. Он участвует в регуляции пигментного обмена в организме. Помимо своего влияния на пигментный обмен этот гормон принимает участие в адаптации животных к цвету
грунта. Секреция МСГ находится под тормозным влиянием гипоталамуса.
Тиреотропный гормон (ТТГ) – гликопротеид,
с молекулярным весом 30000, вырабатывается базофильными клетками. Основная роль этого гормона – стимуляция функции щитовидной железы. Гормон повышает количество внутриклеточного коллоида и стимулирует выделение тироксина. Между гормоном щитовидной железы и ТТГ существуют регуляторные взаимодействия.
Тироксин блокирует реакцию гипофиза на ТТГ – освобождающий фактор и подавляет секрецию ТТГ. Секрецию тиреотропного гормона подавляют адреналин и кортикостероиды. Этим и объясняется снижение активности щитовидной железы при первичных стресс-реакциях, которые вызываются различными факторами, кроме холода. Холод оказывает противоположное
действие – стимулирует выработку тиреотропного гормона.
Андренокортикотропные гормоны – полипептиды, образуются в базо-
фильных клетках. По-видимому, существуют три АКТГ, а не один, как считали ранее. Их молекулярный вес равен 45000 и содержат до 39 аминоксилотных остатков.
АКТГ стимулирует кору надпочечников, особенно пучковой зоны, усиливая секрецию глюкокортикоидов. На выделение минералокортикоидов он не влияет или влияет очень слабо.
Секрецию АКТГ гипофизом в обычных условиях регулирует гипоталамус при помощи релизинг-фактора АКТГ. На выделение этого фактора в свою очередь влияет содержание кортикостероидов в крови. При стрессе секреция АКТГ стимулируется частично нервным путем через гипоталамус, а частично путем возбуждения адреналином центров гипоталамуса и сам гипофиз.
В аденогипофизе синтезируются биологически активные соединения,
регулирующие функцию тимуса, околощитовидной железы и островков Лангерганса. Тимус обеспечивает иммунологическую защиту. Его гормон тимозин стимулирует развитие и поступление в кровь стволовых клеток из костного мозга, а затем их функциональную дифференцировку с образованием Т-
лимфоцитов. Активация островков Лангерганса сопровождается интенсивным синтезом инсулина, глюкагона и липокаина, участвующих в регуляции углеводного и жирового обмена. Под влиянием гипофиза обеспечивается функция околощитовидной железы, которая продуцирует паратгормон, участвующий в регуляции водно-солевого обмена. Считают, что передняя доля
гипофиза вырабатывает до 20 различных гормонов и других биологически активных соединений.
Нейрогипофиз (задняя доля гипофиза). Этот орган нельзя считать эндокринной железой. Он не только образуется как выпячивание дна третьего мозгового желудочка, но и сохраняет связь с головным мозгом в течение всей жизни. В настоящее время точно установлено, что его нейросекреты
синтезируются в супраоптическом и паравентрикулярном ядрах гипоталамуса, а затем переносятся по нервным волокнам в нейрогипофиз, где и выделяются в капилляры. Таким образом, нейрогипофиз – нейросекреторная железа.
Нейрогипофиз выделяет два гормона – антидиуретический гормон (аргинин – вазопрессин) и окситоцин. Эти гормоны относятся к полипептидам.
Содержат 8 аминокислот, 6 из них общие для обоих гормонов.
Антидиуретический гормон (вазопрессин) – АДГ, действует на гладкие мышцы кровеносных сосудов и вызывает их сужение, действует на почки и тормозит диурез. Основное действие АДГ у млекопитающих заключается в стимуляции всасывания воды в дистальных почечных канальцах. Задержка воды в организме находится в прямой зависимости от содержания АДГ в
нервных клетках. Чем больше его, тем сильнее задерживается вода в организме. АДГ оказывает влияние непосредственно на канальцы, особенно на петлю Генле, дистальный извитой каналец и собирательные трубки – он повышает реабсорбцию воды из них и этим самым снижает количество выводимой мочи.
Вазопрессин совместно с другими гормонами надпочечников (минералокортикоиды) и почек (ренин) обеспечивает равновесие электролитов в тканях. Секреция вазопрессина (АДГ) происходит непрерывно и скорость секреции регулируется составом крови и осмотическим давлением. Понижение осмотического давления ослабляет секрецию гормона. Полагают, что рецептором осмотического давления крови служат клетки супраоптического
ядра гипоталамуса. Уменьшение объема циркулирующей крови, охлаждение
организма снижают секрецию АДГ. Накопление продуктов обмена веществ в
крови вызывает раздражение интерорецепторов кровеносных сосудов дуги
аорты и каротидного синуса, а это в свою очередь тормозит продукцию вазопрессина. При недостаточном выделении вазопрессина выделяется большое количество воды с мочой и возникает так называемое несахарное мочеизнурение.
Окситоцин по химической природе пептид. Этот гормон вызывает сокращение гладкой мускулатуры, особенно в стенке матки и в меньшей степени – в стенке мочевого пузыря, желчного пузыря и кишечника. Механизм
действия окситоцина связан с нейтрализацией холинэстеразы – фермента,
который разрушает ацетилхолин. При его повышенной секреции увеличивается количество ацетилхолина. который возбуждает гладкую мускулатуру
матки, что приводит к изгнанию плода (родам). Этот гормон стимулирует также молокоотделение. Окситоцин выделяется во время лактации при раздражении соска. Он влияет также на миоэпителиальные клетки, которые окружают альвеолы молочной железы. Это приводит к поступлению молока в сосок. Он способствует не только сокращению миоэпителиальных клеток
молочной железы, но и стимулирует образование жировых шариков в клетках секреторного эпителия и обеспечивает их выход в полость альвеол.
Окситоцин по своему действию на водный обмен близок к вазопрессину – он тормозит реабсорбцию калия и натрия из первичной мочи и увеличивает фильтрацию в почечных клубочках.
Высокая чувствительность матки к окситоцину свидетельствует о том, что выделение окситоцина в большом количестве в период родов – важный
фактор для нормальной физиологии родов. Эстрогены усиливают действие окситоцина на мускулатуру матки, а прогестерон противодействует ему.
РОЛЬ ГОНАДОТРОПНЫХ И ОВАРИАЛЬНЫХ ГОРМОНОВ В РЕГУЛЯЦИИ ПОЛОВЫХ ЦИКЛОВ У САМОК.
Цикличность половых процессов у самок регулируется сложным нейрогуморальным механизмом. Нервная регуляция осуществляется головным 1 и спинным 8 мозгом, а также проводниковой симпатической 15 и парасимпатической 16 нервной системой. Гуморальная регуляция связана с функцией гипоталамо-гипофиз-овариальной системы и эпифиза. Если перерезать ножку гипофиза, то атрофируются яичники и нарушится половая цикличность. Аналогичные изменения наступают и при перерезке только портальных вен, которые несут кровь от срединного возвышения гипоталамуса к аденогипофизу.
В опытах с пересадкой гипофиза крыс от самцов к самкам и наоборот установлено, что в синтезе гонадотропинов половое различие присуще не гипофизу, а гипоталамусу. Так, гипофиз самцов в организме самок вырабатывает фолликулостимулирующий (ФСГ) и лютеинизирующий (ЛГ) гормоны, и организм функционирует циклично. Гипофиз самок в организме самцов постоянно секретирует только один лютеинизирующий гормон, в результате чего половой цикличности не отмечается. Следовательно, роль гипофиза соподчиненная. Его функция связана с регулирующим влиянием гипоталамуса и других отделов головного мозга. В гипоталамусе клетками ядер
вырабатывается нейросекрет, обеспечивающий гонадотропную функцию гипофиза.
Большая роль отводится нейросекретам переднего гипоталамуса – преоптическому 2, супрахиазматическому 3, паравентрикулярному 4 и супраоптическому 5 ядрам. Супрахиазматическое и преоптическое ядра относятся к
основным ядрам “высшего” центра, регулирующим половые процессы.
Они морфофункционально связаны с таламусом и корой головного мозга. Вырабатываемый ими нейросекрет стимулирует клетки “низшего“ центра, представленного вентромедиальным 6 и аркуатным 7 ядрами.
Клетки “низшего” центра вырабатывают нейросекрет – фактор активации гонадотропной функции – фол- и люлиберины, которые относятся к
низкомолекулярным полипептидам, веществам дистантного (на расстоянии)
действия. В зависимости от его концентрации происходит тоническая (постоянная) или циклическая секреция гонадотропинов.
Аксоны нейросекреторных клеток этих центров анастомозируют между собой и заканчиваются на петлях первичной капиллярной сети 9. К ней подходят также аксоны адреналинергических серотонинергических клеток,
медиаторы которых влияют на синтез и поступление в кровь нейросекрета.
Ядра “низшего” центра побуждают аденогипофиз постоянно секретировать
небольшое количество фол- и люлиберина, необходимых для постоянного выделения ФСГ и ЛГ, то есть такого количества, которое поддерживает аденогипофиз в определенном тонусе (тоническое количество), от которого овуляция не происходит.
Преоптические ядра 2 вырабатывают нейросекрет, стимулирующий функцию клеток “низшего” центра, а он, в свою очередь, активизируют гипофиз на синтез и выведение гонадотропинов в количестве, обеспечивающем овуляцию и развитие желтого тела (овуляторная квота ЛГ). Поэтому “высший” центр иначе называют “овуляторный”.
Механизм функционального взаимодействия гипоталамических ядер в регуляции половых циклов имеет свои особенности. Клетками супрахиазматических ядер 3 вырабатывается серотонин, который тормозит работу клеток
преоптического ядра 2, а следовательно, и угнетает секрецию гонадотропных гормонов гипофиза. При уменьшении концентрации серотонина стимулируются клетки преоптического ядра и повышается гонадотропная функция гипофиза. Таким образом, снижение продукции серотонина в клетках супрахиазматического ядра растормаживает “высший” центр, а следовательно, повышает активность “низшего” центра: увеличивается отдача либерина, активизирующего гонадотропную функцию гипофиза.
Синтез нейросекрета клетками “низшего” центра также возрастает в связи с повышенным выделением дофамина – продукта адреналинергических нейронов гипоталамуса. Дофамин – антагонист серотонина, следовательно, отдача фактора активизации гонадотропной функции определяется отношением дофамина и серотонина.
Аксоны серотонинергических волокон усиливают выработку клетками “низших” центров нейросекрета, способствуют освобождению его из терминалий (окончаний) нейронов и поступлению в кровь первичной капиллярной
сети.
Аксоны серотонинерических волокон, наоборот, тормозят образование и освобождение секрета в терминалях нейронов.
Вазопрессин и окситоцин супраотического 5 и паравентрикулярного 4 ядер, поступив в зону клеток “низшего” центра, активизируют их функцию, в связи с чем в аденогипофизе больше вырабатывается гонадотропинов. Следует отметить, что супраоптические и паравентрикулярные ядра имеют всего
лишь вспомогательное значение в регуляции половых процессов, так как и при разрушении этих ядер гонадотропины синтезируются в колличестве, достаточном для обеспечения циклической функции яичников.