Уровень липидов в крови определяется приемом пищи, деградацией в кишечнике желчными кислотами и липазами, абсорбцией энтероцитами, деградацией в кровеносных сосудах липопротеинлипазами, а также жировыми тканями жировой триглицеридлипазой и гормончувствительными липазами, синтезом de novo в печени и выведение периферическими клетками и печенью[8].
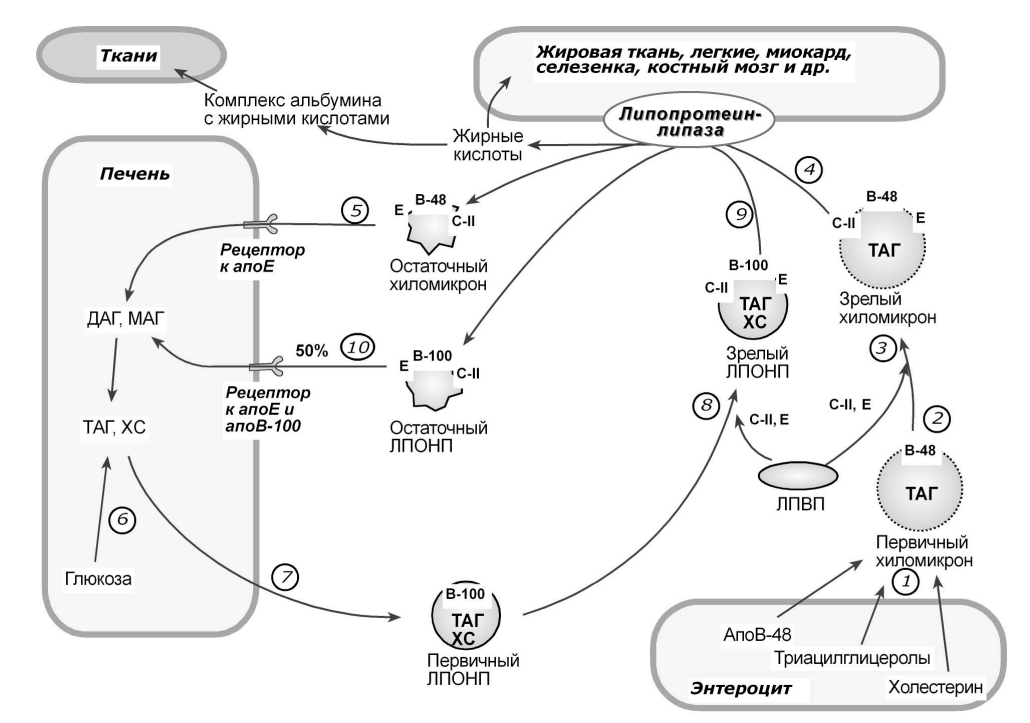
Важно отметить, что липиды переносятся альбуминами и аполипопротеинами в кровь и лимфатическую систему. Липиды и аполипопротеины объединяются с образованием липопротеинов, включая хиломикроны, липопротеины очень низкой плотности (ЛПОНП), ЛПНП, липопротеины промежуточной плотности (ЛПВП) и липопротеины высокой плотности (ЛПВП). Белки-переносчики вырабатываются в печени и кишечнике. Липиды, альбумин и аполипопротеины демонстрируют суточные вариации на животных моделях [80,81] и людях [82].
Кроме того, экспериментальная ночная смена также приводит к повышению уровня триглицеридов в сыворотке крови у здоровых молодых людей [83].
В комплексном обзоре данных о людях Poggiogalle et al. пришли к выводу, что, хотя в большинстве исследований сообщалось о значительных циркадных колебаниях уровней липидов в сыворотке, результаты были противоречивыми в отношении величины изменений и времени пиков и минимумов. Помимо факторов образа жизни, таких как диета и физические упражнения, прием лекарств и сопутствующие заболевания, были отмечены значительные межполовые различия [82].

Центральные часы оказывают существенное влияние на голод, аппетит и прием пищи [84]. СХЯ проецирует сигналы непосредственно в дугообразные, вентромедиальные и дорсомедиальные гипоталамические, паравентрикулярные и латеральные гипоталамические ядра головного мозга, которые являются основными центральными регуляторами энергетического гомеостаза [85,86].
Кишечная абсорбция липидов зависит от перистальтики кишечника, деградации желчными кислотами, энтеральными липазами и транспортерами, экспрессируемыми на энтероцитах.
Моторика желудочно-кишечного тракта [92] и выработка желчных кислот и липазы поджелудочной железы демонстрируют циркадные закономерности [93].
CD36 и белки-переносчики жирных кислот участвуют в абсорбции свободных жирных кислот (СЖК) и моноацилглицерина, тогда как CD36 и C1-подобный белок Нимана-Пика (NPC1LP) участвуют в поглощении свободного холестерина [8,97]. Кроме того, АТФ-связывающие кассетосвязывающие белки G5 (ABCG5) и G8 (ABCG6) экспрессируются в апикальных мембранах энтероцитов и отвечают за отток свободного холестерина в кишечник [97]. Мыши, экспрессирующие мутантный белок CLOCK∆19/∆19, имеют более высокие уровни CD36 и NPC1LP [98]. Кроме того, у мышей с нокаутом BMAL1 наблюдалось увеличение экспрессии NPC1LP и снижение экспрессии ABCG5 и 8 [99].
Это образование опосредовано микросомальным белком-переносчиком триглицеридов (MTP). У мышей с нокаутом CLOCK наблюдалась повышенная экспрессия MTP, что указывает на прямой циркадный контроль MTP [100]. Хиломикроны подвергаются дальнейшей модификации за счет приема аполипопротеина A-IV, который также находится под прямым влиянием BMAL1 [101] и затем высвобождается в лимфатическую систему. Свободные жирные кислоты высвобождаются из хиломикронов под действием липопротеинлипазы. Повышенная экспрессия липопротеинлипазы была показана у мышей с нокаутом REV-ERBα, а CLOCK дополнительно усиливал ее экспрессию, что указывает на прямой контроль со стороны гена CLOCK [102]. Активность липопротеинлипазы также может косвенно регулироваться молекулярными часами. Активируемый пролифератором рецептор γ (PPARγ), важный активатор липопротеинлипазы [103], был снижен в клетках с дефицитом BMAL1 [104]. В соответствии с этим сообщалось о циркадных изменениях липопротеинлипазы [105]. Остаточные хиломикроны улавливаются печенью с помощью рецепторов ЛПНП (ЛПНП). Ма и др. сообщили, что удаление BMAL1 привело к снижению экспрессии LDLR [106]. Многие данные свидетельствуют о циркадном контроле синтеза липидов в печени. Связывание регуляторного элемента стерола
β-Гидрокси-β-метилглутарил-КоА (HMG-CoA) редуктаза, важный фермент синтеза холестерина и основная мишень терапии статинами, демонстрирует суточную ритмичность с самой высокой экспрессией в полночь [109,110].
Холестерин высвобождается через желчные кислоты, которые, как упоминалось выше, находятся под контролем молекулярных часов [96]. Ферменты, участвующие в синтезе триглицеридов в печени, также демонстрируют циркадный контроль, как показано на мышах с нокаутом PER [112]. У мышей с дефицитом BMAL1 наблюдалось более высокое производство ЛПОНП и повышенные концентрации ЛПОНП в печени [99]. MTP играет центральную роль в формировании ЛПОНП, поскольку он связывает апоВ-100 с ЛПОНП [8]. Мыши с дефицитом BMAL1 [99] и CLOCK [98,100] показали более высокую экспрессию MTP. Увеличение концентрации ЛПОНП также может быть связано с увеличением продукции аполипопротеина A-IV в печени на модели мышей с дефицитом BMAL1 [101]. Циркулирующие частицы ЛПОНП разлагаются до ЛПНП и ЛПНП посредством липопротеинов и печеночных липаз, которые, как сообщается, демонстрируют циркадный ритм [105]. Свободные жирные кислоты и глицерин также высвобождаются из жировой ткани.
Гормоночувствительная липаза блокируется инсулином, что также оказывает сильное циркадное влияние [115]. Повышенное высвобождение СЖК из жировой ткани у мышей с дефицитом BMAL1 приводит к образованию эктопической жировой ткани в печени и скелетных мышцах [116]. Частицы ЛПНП являются основным переносчиком холестерина. Если его поглощение задерживается периферическими клетками или печенью, он претерпевает модификации, такие как окисление активными формами кислорода, что приводит к образованию атерогенных частиц оксЛПНП [8]. Поглощение ЛПНП печенью обусловлено печеночным ЛПНП, который оказывает циркадное влияние [106]. Циркулирующие частицы ЛПНП диффундируют через эндотелиальный барьер и захватываются сосудистыми макрофагами через рецепторы-мусорщики. У мышей, экспрессирующих мутантный белок CLOCK∆19/∆19, наблюдалась повышенная экспрессия CD36 и рецептора-мусорщика A1, что способствовало развитию атеросклероза [98]. АТФ-связывающая кассета А1 (ABCA1) является важной составляющей ЛПВП и отвечает за обратный транспорт холестерина [8]. У мышей, экспрессирующих мутантный белок CLOCK∆19/∆19, наблюдалась сниженная экспрессия ABCA1 [98]. SCN влияет на выработку кортизола посредством проецирования афферентов на паравентрикулярное ядро [117].
Проекции SCN также влияют на выработку мелатонина [4], который стимулирует секрецию инсулина [119]. Инсулин стимулирует липопротеинлипазу, способствуя притоку свободных жирных кислот к жировой ткани и блокируя там липолиз [8].
Наконец, SCN проецирует афференты на гипофиз, влияя на выработку гормона роста [120].
Гормон роста облегчает липолиз в жировой ткани [121].
Следует отметить, что на выработку гормона роста также напрямую влияет сон, поскольку он в основном вырабатывается во время глубокого (N3) сна [122].