Общей (пожалуй, кроме вельвичии) особенностью гнетовых является склонность к «быстрому сексу»: репродуктивные процессы протекают стремительно. Впрочем, это смотря с чем сравнивать. У цветковых время от опыления до оплодотворения измеряется часами (рекорд – 12 минут). У большинства голосеменных – неделями и месяцами. Гнетовые занимают явно промежуточное положение. У гнетума это 6-8 дней, у эфедры 10-36 часов. В последнем случае такой короткий интервал связан с сокращением расстояния до архегония и быстрым прорастанием пыльцы (1-2 часа). Вообще у гнетовых пыльца прорастает почти так же быстро, как у цветковых. Это в равной мере относится и ко многим другим репродуктивным процессам.
Развитие женского гаметофита и оплодотворение происходит принципиально по-разному у разных родов гнетовых. Их невозможно заподозрить в сколько-нибудь близком родстве, если смотреть только на эту группу признаков. Гнетовые в этой части представляют собой настоящее «божье хобби»: творец почему-то избрал объектом именно их и дал простор фантазии.
ЭФЕДРА ушла от хвойных не очень далеко.
Ссылка на источник изображения
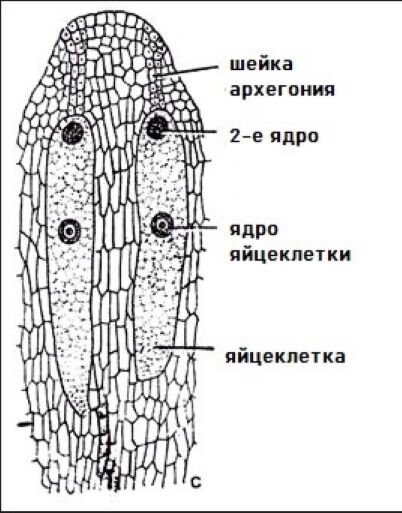
Из мегаспоры через обычную свободноядерную стадию у нее развивается массивный женский (мега-) гаметофит из обычных гаплоидных одноядерных клеток. В нем образуются опять же обычные архегонии в количестве двух. Они показаны на рисунке. Что у них не вполне обычно, так это шейка. В ней десятки клеток. Это самая мощная шейка архегония среди голосеменных. Необычным является поведение центральной клетки архегония. Её ядро делится, но клеточная стенка не образуется. Клетка остается 2-ядерной: это ядро яйцеклетки и еще одно ядро.
К тому времени, когда пыльца посредством опылительной капли попадает на мегаспорангий, в нем уже имеется глубокая (самая глубокая среди голосеменных) воронкообразная пыльцевая камера. Пыльцевое зерно оказывается прямо на поверхности женского гаметофита возле шейки архегония. Пыльцевая трубка вообще не касается клеток мегаспорангия (нуцеллуса). Она совсем короткая, простая, и растет исключительно в специально предназначенной для этого шейке архегония. Пыльцевая трубка доставляет 2 спермо-ядра в единой цитоплазме прямиком к той самой 2-ядерной клетке. Наклевывается формат 2 + 2. Реализуется ли он?
Реализуется! В результате получается как бы две зиготы: основная (от слияния спермо-ядра с ядром яйцеклетки) и дополнительная. Из основной зиготы развивается зародыш. Вторая тоже делится. Из нее развивается некая многоклеточная диплоидная структура, которая предположительно (это пока не доказано) имеет значение для питания зародыша на ранних стадиях его развития. Это настоящее двойное оплодотворение, которое в прежние времена считалось важнейшим аргументом близости гнетовых к цветковым. Теперь уже не считается. У примитивных цветковых нет характерного для остальных цветковых двойного оплодотворения с образованием триплоидного эндосперма. Значит, последнее возникло уже после того, как предки современных цветковых отделились от предков современных голосеменных. Значит, двойное оплодотворение эфедры возникло независимо от двойного оплодотворения цветковых. Независимо и значительно раньше. В стремлении найти применение второму спермию природа сделала две попытки: первую пробную у эфедры и вторую очень эффективную у цветковых.
ГНЕТУМ
В части женского гаметофита и оплодотворения представляет собой совершенно уникальный случай. Странности начинаются с самого начала. У всех семенных растений, включая эфедру, диплоидная археспориальная клетка в мегаспорангии подвергается редукционному делению (мейозу), в результате чего формируются 4 гаплоидных клетки, из которых лишь одна становится мегаспорой. Из нее формируется МОНОСПОРИЧЕСКИЙ женский гаметофит. У гнетума после мейоза образуется большая 4-ядерная «мега-мега» спора. Из нее формируется ПОЛИСПОРИЧЕСКИЙ женский гаметофит.
Его развитие, как и у остальных голосеменных, начинается со стадии свободных ядер: ядра делятся, но клеточные стенки не образуются. За 8 раундов клеточных делений из исходных 4 ядер образуется до 1000 ядер.
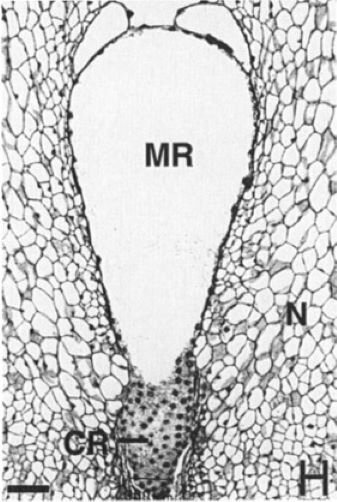
N – нуцеллус (мегаспорангий). Он имеет клеточную структуру. В центре располагается женский гаметофит. В нижней части снимка (CR) показана та его часть, где началась целлюрялизация – образование клеточных стенок. Большая часть женского гаметофита – это одна огромная клетка с центральной вакуолью (белое поле) и постенной цитоплазмой, в которой находятся сотни ядер. Слой цитоплазмы очень тонкий. Поэтому на снимке видны только ядра как черные точки. Тут бы самое время завершить целлюрялизацию всего женского гаметофита, сформировать архегонии, в архегониях – яйцеклетки, подготовить их к оплодотворению. Поразительно, но факт: у гнетума ничего этого не происходит. На представленной фотографии мы видим женский гаметофит в состоянии половой зрелости, т.е. полностью готовый к оплодотворению!
(А) У голосеменных, включая эфедру, женский гаметофит большой, включает тысячи клеток, в том числе, пару яйцеклеток (egg cells) по одной в каждом архегонии. (С) У цветковых женский гаметофит (зародышевый мешок) очень маленький (по линейным размерам в десятки, по объему – в сотни раз меньше, чем у голосеменных), строго 7-клеточный с единственной яйцеклеткой. (B) У гнетума женский гаметофит примерно в 10 раз меньше, чем у остальных голосеменных (промежуточный по размеру между голосеменными и цветковыми). Большую часть его занимает центральная вакуоль и постенная цитоплазма с сотнями свободных ядер.
Именно в эту гигантскую многоядерную клетку у гнетума врастает пыльцевая трубка. Ее оболочка рассасывается, высвобождаются два спермо-ядра. Перед спермиями встает вопрос: кого будем оплодотворять? Ведь яйцеклеток-то нет: какое ядро оплодотворишь, то и яйцеклетка! Дорогой читатель мужского пола, представьте, что вы с другом попали в изолированное сообщество близнецов женского пола, которые «все равны как на подбор», все «в самом соку», и все – «не против». Сразу вспоминается буриданов осел: он не смог выбрать даже из двух одинаковых вариантов. А тут – сотни! Чем руководствуются несчастные спермо-ядра в своем выборе, науке пока не известно. Однако оплодотворение так или иначе происходит. Из зиготы развивается зародыш. Изредка их бывает два, но не больше.
Сразу после оплодотворения начинается целлюрязация женского гаметофита. Казалось бы, это совсем просто: ядра разгораживаются клеточными стенками и дело получается с концом. Но гнетовые не были бы гнетовыми, если бы действовали по-простому.
Фотографии сделаны в специальной технике, которая позволяет видеть только ядра. На первой – конец свободноядерной стадии (FN = free nucleus = свободные ядра). На второй видно, как ядра начинают группироваться по несколько штук. Вокруг этих группировок и формируются клеточные стенки (MNC = multinucleate cell = многоядерные клетки). Затем несколько ядер, оказавшихся в одной клетке, сливаются. Получается одноядерная полиплоидная клетка. Это изображено на третьей фотографии (UNC = uninucleate cell = одноядерные клетки). Из женского гаметофита у гнетума, как и у всех голосеменных, формируется эндосперм – питательная ткань семени. Остальные голосеменные имеют гаплоидный эндосперм. Считается, что это «минус». Большинство цветковых имеют триплоидный эндосперм. Считается, что это жирный «плюс». У гнетума эндосперм полиплоидный!!!
У саговниковых и гинкговых к моменту оплодотворения женский гаметофит достигает почти окончательного размера. У хвойных подрастает после оплодотворения, но совсем немножко. Это не эффективно: зародыш то ли будет, то ли нет, а вещество и энергия расходуются. Полиплоидный супер-эндосперм гнетума показывает свои супер-способности сразу же после образования. Он растет, как на дрожжах. Его размер увеличивается не меньше, чем в 1000 раз! Причем, растет он по принципу цветковых растений: только после образования полноценного зародыша. Поэтому эффективность использования вещества и энергии получается максимальная. Для чего как раз и затевался описанный выше эксперимент природы. В эволюционной теории это явление называется гетерохрония: изменение времени закладки и темпа развития органов у потомков по сравнению с предками. В данном случае она выразилась в том, что женский гаметофит достиг стадии половой зрелости на том этапе своего «вегетативного» развития, на которой женские гаметофиты других голосеменных еще «ходят пешком под стол».
ВЕЛЬВИЧИЯ
Интересно, что же в этой области «учудила» вельвичия. Ведь не могло же самое загадочное растение нашей планеты остаться в стороне от общего для гнетовых процесса «перманентной революции». Начнем с того, что многие новообразования, которые мы рассмотрели на примере гнетума, есть и у вельвичии: полиспорический женский гаметофит, отсутствие архегониев и конкретных яйцеклеток, полиплоидный эндосперм. Следовательно, эти достижения были уже у их общего предка.
Со времени открытия вельвичии к ней в течение полутора веков было приковано внимание всего ботанического сообщества. Ученые изучали ее наперегонки. В погоне за сенсацией это нередко приводило к ошибкам. "Qui quaerit, reperit": говорили древние. Вот и у вельвичии находили то, чего у нее нет: моноспорический женский гаметофит, двойное оплодотворение. Эти открытия потом приходилось «закрывать». Что же реально нового и интересного есть у вельвичии в развитии женского гаметофита и оплодотворении?
Свободноядерная стадия развития женского гаметофита есть у всех голосеменных, но у всех она в это время состоит из центральной вакуоли и постенной цитоплазмы (мы это только что видели на примере гнетума). У вельвичии никакой центральной вакуоли нет. Есть плотная цитоплазма, в которой равномерно распределены ядра.
Картинки, как видите, взяты из статьи того же автора, который сделал только что рассмотренную нами публикацию по гнетуму. Только от первой статьи до второй прошло 17 лет. За это время микроскопическая, фотографическая и полиграфическая техника заметно продвинулись. Поэтому микрофотографии выглядят ярче, особенно если препараты окрашены «под Гжель». На нижней фотографии изображено то же, что и на верхней, только под большим увеличением. Там хорошо видна агрегация ядер в группы перед целлюляризацией и последующим слиянием нескольких гаплоидных ядер в одно полиплоидное. Только уровень плоидности будущего эндосперма у вельвичии получается заметно выше, чем у гнетума: в одно полиплоидное ядро, бывает, сливаются до 12 гаплоидных ядер. В то же время, некоторые гаплоидные ядра ни с кем не сливаются. Одно из них впоследствии выступит в роли яйцеклетки, т.е. сольётся со спермо-ядром.
Ее обнаружил уже первоописатель вельвичии Джозеф Гукер в 1862 г. С тех пор ее многократно описывали под разными названиями. Суть в том, что дюжина или больше многоядерных клеток на микропилярном конце женского гаметофита инициируют его трубчатое расширение по направлению к растущей от микропиле пыльцевой трубке. Красной стрелкой обозначена зона делящихся клеток на конце трубки, ее точка роста. Проталлиальная трубка несёт яйцеклетку к спермиям. Это уникальный для голосеменных, редчайший для семенных (что-то похожее есть у очень немногих цветковых) растений случай, да и вообще редкое явление в «большом» (не только растительном) мире. Принцесса обычно сидит на попе ровно и ждет принца, который прорывается к ней через огонь, воду и медные трубы. Что это, если не сексизм? В данном случае налицо полное равноправие: женское и мужское начало, Инь и Ян, движутся навстречу друг другу, проходят каждое ровно половину пути.
В качестве яйцеклетки у вельвичии выступает одно из свободных гаплоидных ядер. Достоверных фактов двойного оплодотворения не выявлено. Скорее всего, вельвичия утратила эту способность, присутствовавшую у ее предков. Скорее всего, потому, что от нее не было никакой реальной пользы.
Тему формирования зародыша мы, пожалуй, пропустим как не особенно актуальную. Тем более, что между родами тут нет больших различий. А вот на семенах в целом и способах их распространения – остановимся. Но не здесь. Дело в том, что по этим признакам значительные различия есть не только между родами, но и внутри них. Обсуждая какую-то таксономическую группу растений, мы обычно сначала даем ее общую характеристику, затем переходим к описанию внутреннего разнообразия. С гнетовыми мы изменили этому принципу потому, что различий между тремя родами было многократно больше, чем сходства. Поэтому в последнем разделе общей для каждого рода информации мы не будем уделять много внимания, зато сосредоточимся на внутриродовой систематике и различиях между видами. Сначала рассмотрим эфедру как самый древний род, затем два других.