Обсуждая голосеменные в целом, мы упоминали два типа строения ствола (стебля): (1) пикноксильное (от греч. pyknos - плотный и xylon - древесина), когда он состоит, в основном, из древесины; (2) маноксильное (от греч. manos - рыхлый, неплотный), когда древесины мало, много коры и сердцевины. Нормальные голосеменные подклассы или целиком пиноксильные (хвойные, гинкговые) или целиком маноксильные (саговниковые). Но у гнетовых всё не как у всех. У них налицо большое разнообразие.
Гнетовые, по современным представлениям, ближе всего к хвойным или даже находятся внутри этого подкласса. Поэтому вполне естественно, что ближайший к предкам род эфедра имеет примерно такое же, как у хвойных, строение стебля, т.е. пикноксильное. В молодом, 1-летнем стебле камбий и производные от него проводящие ткани, располагаются отдельными пучками. Потом они сливаются в сплошное кольцо. В холодный и/или сухой сезон камбий отдыхает, в теплый и/или влажный сезон – работает, образует новый слой луба и древесины.
Эфедра
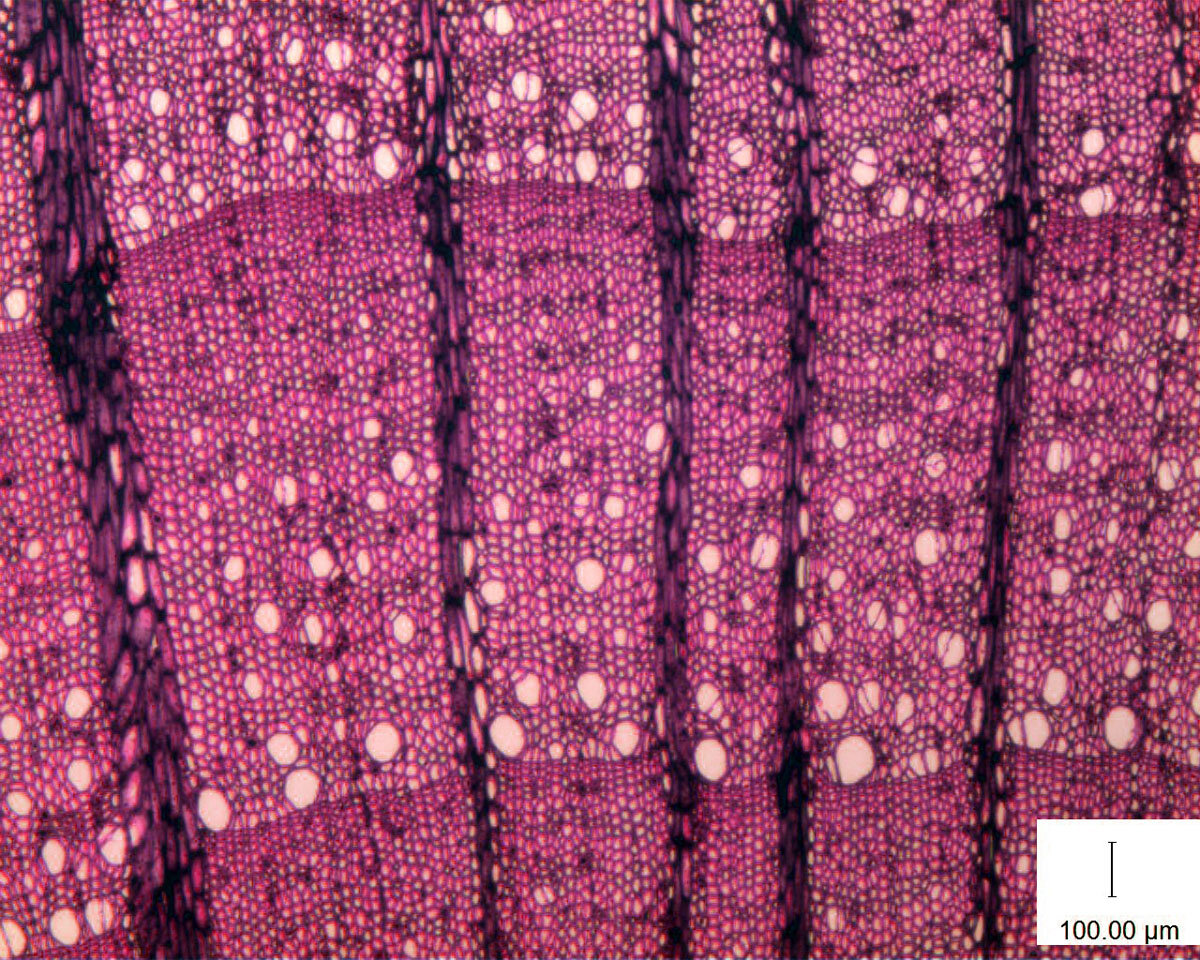
Как видите, они устроены так же, как у наших хвойных: ранняя древесина с широкими трахеидами, поздняя древесина с узкими трахеидами. Клетки с огромными просветами – те самые сосуды, которые мы обозначили как первый характерный и общий для всех гнетовых признак, отличающий их от голосеменных и сближающий их с цветковыми. В ранней древесине их много и они крупнее. По направлению к поздней древесине их всё меньше, меньше и их размер. Это был калифорнийский вид Ephedra trifurca.
А теперь посмотрим на вельвичию
Ссылка на источник изображения
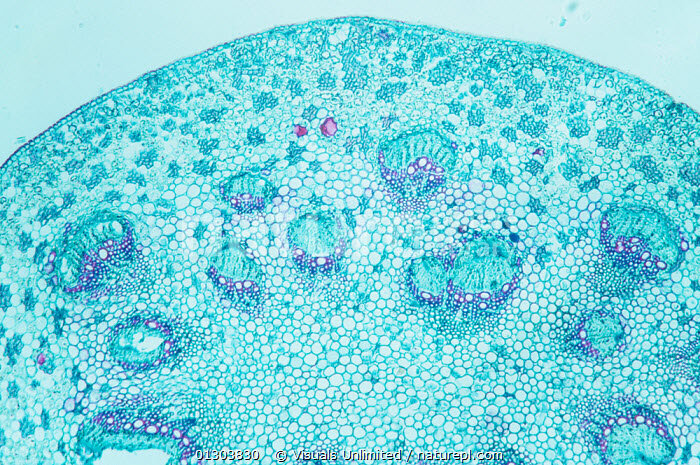
Живые клетки окрашены в голубой цвет, мертвые – в фиолетовый. Как видите, нет ничего общего с эфедрой. Структура чисто маноксильная: очень много живой паренхимной ткани, сердцевина смыкается с корой. Примерно между ними широким кольцом располагаются проводящие пучки. Голубая их половинка – это живые клетки луба, фиолетовая – мертвые клетки древесины: мелкие – трахеиды, крупные – сосуды. Древесины совсем мало. На поперечном срезе она занимает несколько процентов от его общей площади. Вельвичия, несомненно, произошла от пикноксильных, общих с хвойными и эфедрой, предков. Значит, ее маноксильность – это вторичный признак, возможно, адаптация к жизни в пустыне (в паренхимных клетках сердцевины и коры запасается вода).
У вельвичии, в отличие от эфедры и гнетума, нет механизма вторичного утолщения ствола. Его высота с возрастом медленно увеличивается (до 0,5 м) за счет первичного роста. Постепенно увеличивается также и диаметр каждого следующего яруса. А диаметр основания почти не меняется. Поэтому ствол старого растения имеет форму перевернутого конуса.
Ссылка на источник изображения
Теперь рассмотрим гнетум
Ссылка на источник изображения
A – это рисунок среза с натуры. На нем, как будто бы видны годичные кольца древесины. Нет, эти кольца не годичные и состоят не только из древесины. Как они образовались, показано на схеме (B). В начале жизни данного фрагмента ствола он развивался точно так же, как ствол эфедры или любого представителя хвойных. Камбий образовал проводящие пучки, к концу данного цикла роста они почти сомкнулись. Тут бы камбию спокойно заснуть, чтобы проснуться в начале следующего цикла роста побегов и образовать следующее кольцо древесины. Он и заснул, но … не проснулся. В начале следующего цикла роста камбий образовался de novo из живых клеток коры, причем не сплошным кольцом, а снова отдельными фрагментами. Из них образовалось новое кольцо разрозненных проводящих пучков. Потом этот цикл погибели старого и инициации нового камбия происходил еще два раза (всего на срезе представлено 4 таких ростовых цикла). Эти циклы (соответственно, кольца пучков) редко бывают годичными, чаще за год происходит два цикла роста, соответственно, образуется два кольца пучков.
Маноксильная это структура или пикноксильная? Трудно сказать: это нечто среднее. Такая модель развития тоже сформировалась из пикноксильной. Это предположительно адаптация к существованию в жизненной форме лианы: возможно, она повышает гибкость ствола (у двух древовидных видов гнетума ствол обычный, как у хвойных и эфедры). Кстати, обратите внимание на резкую асимметрию колец: на одном радиусе они многократно шире, чем на другом. Такое явление характерно для горизонтально ориентированных стволов любых лиан: на нижней стороне утолщение происходит в разы активнее.
На предыдущей картинке не показана клеточная структура проводящих пучков гнетума, поэтому непонятно, есть ли там сосуды. Конечно, есть.
Сосуды как более или менее сквозные широкие трубки для проведения воды от корней к листьям – это универсальная и очень эффективная замена трахеидам в эволюции нескольких групп высших растений. С одной стороны, они есть даже у некоторых папоротников, с другой стороны, их нет даже у некоторых цветковых. Это означает, что они несколько раз независимо возникали в ходе эволюции. Самые совершенные и самые эффективные сосуды, конечно, у цветковых. Они так далеко ушли от исходной трахеиды, что вообще перестали быть похожими на нее: их диаметр достигает 500 мкм, причем, длина членика в некоторых случаях равна его диаметру.
Из голосеменных сосуды есть лишь у гнетовых. От исходной трахеиды они ушли не так далеко, но для голосеменных это большое достижение. Приведем таблицу, которая основана на трудах крупнейшего специалиста в области древесины гнетовых Шервина Карлквиста (Sherwin Carlquist) из его уже упомянутой выше работы (Carlquist, 1996). Самые узкие сосуды у вельвичии. Оно и понятно. В пустыне Намиб с водой «напряженка». Ее надо беречь, запасать, а не гонять туда-сюда по проводящим тканям. У эфедры сосуды немногом крупней, чем у вельвичии, видимо, по той же причине. А вот у гнетума, как и у любой лианы, сосуды мощные, сравнимые по диаметру с сосудами многих цветковых, конечно, не лиан. У лиан из числа покрытосеменных они еще шире.
Листья гнетовых, как и большинство признаков, на удивление разные у трех родов. Разные настолько, что говорить о них в целом очень сложно. Это потому, что растения живут в разных климатических условиях, притом еще и адаптируются к ним по-разному. Однако они не всегда были такими. Ведь это монофилетическая группа растений. Значит, у них был общий предок. В последнее время описано много разного рода окаменелостей, относящихся к гнетовым. Поэтому появилась возможность проследить их эволюцию.
Перекрестно-парное листорасположение – очень характерный для гнетовых признак. У современных хвойных перекрестно-парное листорасположение тоже встречается, но только в относительно продвинутых родах, а общий предок гнетовых и хвойных явно имел спиральное листорасположение. Значит, для гнетовых это явное новообразование.
Различаются по размеру, форме, жилкованию. У их общего предка листья, похоже, были условно средними: не большими и не маленькими, не узкими и не широкими.
Современные виды эфедры дифференцировались в олигоцене, когда произошло резкое похолодание климата, он стал более сухим. Чтобы соответствовать ему, у эфедры сформировались крайне редуцированные, мелкие, линейные листья. Они основаниями образуют обкладку, окружающую узел и базальную часть междоузлия. У каждого листа 2, редко 3, параллельные жилки.
Гнетум оказался в дождевых тропических лесах. Поэтому листья у него стали как у двудольных: среднего размера, широкие с перистым или сетчатым жилкованием.
Листья вельвичии – это отдельная история. Мы ее уже по большей части рассказали. Листья вельвичии длиннющие, широкие, лентовидные. На каждом листе много относительно крупных параллельных продольных жилок, которые соединены относительно мелкими поперечными жилками.
С гнетумом всё ясно: у него листья такие же, как у большинства других лиан дождевого тропического леса. А вот эфедра и вельвичия удивляют. Та и другая живут в аридных условиях, но листья у них совершенно разные. Листья эфедры, как правило, вообще никакие. Они мельчайшие и едва зеленые, почти без устьиц. У большинства видов эфемерные: живут лишь до тех пор, пока не вырос и не сформировался стебель, потом опадают. Зато стебель всегда интенсивно зеленый, в его эпидерме много устьиц. Он, собственно, и фотосинтезирует.
Максимальная редукция листьев или их утолщение – обычное дело для пустынных растений. Вельвичия совершенно не вписывается в эту картину. Площадь одного ее листа достигает 10 кв. м и он совершенно плоский. Вот так ксерофит! Как же она живет в пустыне с такими листьями? Оказываются, они не просто не мешают, но активно помогают вельвичии выжить. Главным источником влаги в пустыне Намиб является туман, который окутывает побережье 300 дней в году. Конденсируясь на огромных листьях вельвичии, влага поглощается через устьица и поступает в проводящую систему растения. Поэтому неудивительно, что вельвичия обладает исключительно большим числом устьиц на обеих сторонах листа (до 22 000 устьиц на 1 кв. см!). Устьица погруженные, но не так глубоко, как это обычно бывает у других ксерофитов.
Это были вегетативные органы гнетовых. В следующий раз обсудим их репродуктивные структуры.