Повышенная продукция супероксидных анионов приводит к окислительному стрессу, который связан с рядом сердечно-сосудистых заболеваний, включая атеросклероз (АС), гиперхолестеринемию, гипертонию, диабетическую кардиомиопатию, диабетическую ретинопатию и ишемическую болезнь сердца.
1.Anea CB, Cheng B, Sharma S, Kumar S, Caldwell RW, Yao L, Ali MI, et al. Increased superoxide and endothelial NO synthase uncoupling in blood vessels of Bmal1-knockout mice. Circ Res. . 2012;111:1157–1165. doi: 10.1161/CIRCRESAHA.111.261750. [DOI] [PMC free article] [PubMed] [Google Scholar]
Эпидемиологические данные свидетельствуют о том, что гомоцистеин (Hcy) является независимым фактором риска развития АС.
2.Liu W, Redmond EM, Morrow D, Cullen JP. Differential effects of daily-moderate versus weekend-binge alcohol consumption on atherosclerotic plaque development in mice. Atherosclerosis. 2011;219:448–454. doi:10.1016/j.atherosclerosis.2011.08.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
Эпидемиологические исследования показали, что слегка повышенный уровень ГОМОЦИСТЕИНА в плазме является независимым фактором риска развития АС [13].
ГОМОЦИСТЕИН (Hcy) — это небелковая, серосодержащая аминокислота, которая образуется исключительно путем деметилирования метионина [14].
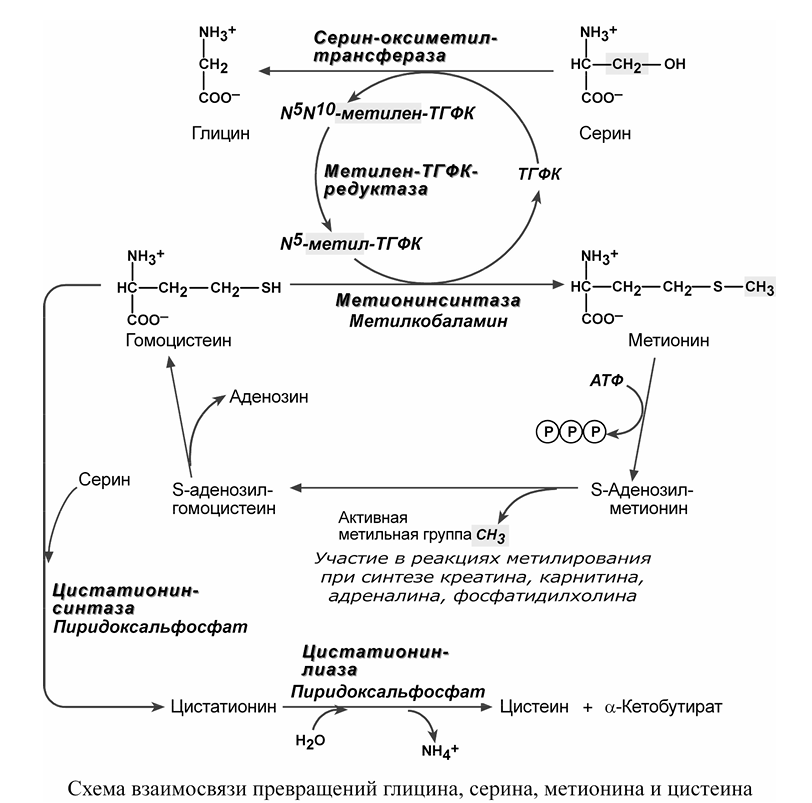
гомоцистеин является предшественником S-аденозилметионина (SAM), первичного донора метильной группы для большинства биологических метилирований, в частности метилирования ДНК, которое является формой эпигенетической регуляции генов, которая функционирует в сочетании с измененными профилями связывания факторов транскрипции [15].
Вместе эти модификации часто приводят к подавлению экспрессии генов, когда они происходят в регуляторных геномных регионах, особенно в промоторных регионах [16].
16.Yoon HY, Kim YW, Kang HW, Kim WT, Yun SJ, Lee SC, Kim WJ, et al. DNA methylation of GSTP1 in human prostate tissues: pyrosequencing analysis . Korean J Urol. . 2012;53:200–205. doi: 10.4111/kju.2012.53.3.200. [DOI] [PMC free article] [PubMed] [Google Scholar]
Метилирование- эпигеномный механизм участвующий в неоплазии клеток рака предстательной железы, глутатионтрансфераза также зависима от механизмов метилирования и деметилирования
Сообщалось, что ген белка, связывающего жирные кислоты, стал гипометилированным, а экспрессия мРНК увеличилась в культивируемых макрофагах и пенистых клетках, обработанных Hcy [17].
В цикле реакции переноса метильной группы ГОМОЦИСТЕИН (Hcy) реметилируется в метионин, затем S-аденозилгомоцистеин (SAH) высвобождается как побочный продукт, и этот цикл поддерживается ДНК-метилтрансферазами (DNMT), включая DNMT3 и DNMT1 [18].
Большинство исследований в области метилирования ДНК относятся к биологии развития, биологии рака и исследованиям с целевыми делециями мышиного DNMT1, который является «поддерживающей» метилтрансферазой, которая обеспечивает правильную передачу профиля метилирования от материнских к дочерним клеткам во время деления клеток [19].
19.Morawietz H. Acromegaly and oxidative stress: impact on endothelial dysfunction and atherosclerosis. Horm Metab Res. . 2013;45:255–256. doi: 10.1055/s-0033-1341495. [DOI] [PubMed] [Google Scholar]
В последние годы исследователи сосредоточились на роли метилирования ДНК при АС, пытаясь получить новые сведения о причине АС и лучше понять эпигенетические механизмы, лежащие в основе патогенеза АС [20]. Laukkanen et al. [21] исследовали метилирование CpG-островков, расположенных на кодирующей последовательности гена EC-SOD в поражениях АС у гиперлипидемических кроликов.
Однако связь между метилированием ДНК и экспрессией EC-SOD не была подробно изучена, в частности механизмы, посредством которых метилирование ДНК регулирует экспрессию EC-SOD и нарушает баланс между окислением и антиокислением при развитии АС.
Материалы и методы
Всего 36 самцов мышей C57BL/6J ApoE –/ – (возраст 6 недель; 25 г) были получены из Центра животных Пекинского университета медицинских наук (Пекин, Китай).
Они содержались по отдельности в комнате с контролируемым климатом при температуре 24 °C и влажности 60 %.
Всего 12 самцов мышей C57BL/6J (возраст 6 недель; 20–25 г; Центр животных Пекинского университета медицинских наук) кормили обычным диетами для мышей (Joint Feed Co., Ltd, Пекин, Китай) и использовалась в качестве нормальной контрольной группы (N-контроль).
напомню про АроЕ+\-
3 группы мышей
1группа
36 мышей C57BL/6J ApoE –/ – были разделены на три группы (n=12/группа) и содержались в течение 15 недель на следующих диетах:
(1) контрольная группа мышей ApoE –/ – (A-контроль), питавшаяся обычной диетой для мышей;
(2) группа Meth, питавшаяся обычной диетой плюс 1,7% метионина для установления модели гипергомоцистеинемии (HHcy);
(3) группа Meth-F, питавшаяся обычной диетой плюс 1,7% метионина (вес/вес), 0,006% фолиевой кислоты и 0,0004% витамина B 12 (Weitonglihua, Пекин, Китай).
Все мыши имели свободный доступ к пище и воде.
Лечение лабораторных животных и используемый экспериментальный протокол следовали рекомендациям, установленным Нинсяским медицинским университетом (Иньчуань, Китай), и были одобрены Комитетом по уходу за лабораторными животными Нинсяского медицинского университета.
Утром последнего дня периода диеты мышей в каждой группе анестезировали 20% этилкарбаматом (1000 мг/кг) путем внутрибрюшинной инъекции, кровь собирали путем сердечной пункции, и мышей усыпляли путем обескровливания.
Сыворотку отделяли центрифугированием (1000 g в течение 10 мин при 4 °C), и все образцы хранили при температуре –80 °C до дальнейшего анализа. Мышей группировали и лечили, как описано в нашем предыдущем исследовании [17].
Определение уровней ГОМОЦИСТЕИНА и липидов в сыворотке у мышей Концентрации Hcy, общего холестерина (ОХ), триглицеридов (ТГ), липопротеинов высокой плотности (ЛПВП) и липопротеинов низкой плотности (ЛПНП) определяли с помощью системы ADVIA 2400 Chemistry System (Siemens AG, Мюнхен, Германия) с помощью обычных ферментативных методов [17].
Результаты Повышенный уровень супероксидного аниона способствует образованию атерогенных поражений, вызванных HHcy
Чтобы оценить атерогенные эффекты HHcy у мышей ApoE –/–, которых кормили пищей с высоким содержанием метионина, атерогенные поражения исследовали с помощью H&E (Рисунок 1A) и окрашивания масляным красным O (Рисунок 1B).
Результаты показали, что площадь атерогенных поражений была значительно увеличена у мышей ApoE –/–, которых кормили пищей с высоким содержанием метионина (Рисунок 1B),
в то время как площадь атерогенных поражений была уменьшена у мышей ApoE –/– после того, как мышей кормили пищей с высоким содержанием метионина, фолиевой кислоты и витамина B 12.
Концентрация Hcy была увеличена в группе Meth и снижена, когда мышей ApoE –/– кормили пищей с высоким содержанием метионина, фолиевой кислоты и витамина B 12 (Рисунок 1C).
Кроме того, было также обнаружено, что концентрация Hcy в сыворотке положительно коррелировала с областями атеросклеротического поражения (r=0,6862; P=0,0016; Рисунок 1D).
Кроме того, уровни TG, TC и LDL были повышены у мышей ApoE –/–, которых кормили диетой с высоким содержанием метионина, тогда как содержание липидов было подавлено, когда мышей кормили фолатом и витамином B12 ( Рисунок 1E).
Однако концентрации HDL были снижены в группах с диетой с высоким содержанием метионина и повышены после того, как мышей кормили диетой с фолатом и витамином B12 ( Рисунок 1E), что подразумевает, что диета с высоким содержанием метионина может вызывать HHcy у мышей ApoE –/–, тогда как фолат и витамин B12 способны смягчать эффекты HHcy.
Окислительный стресс играет решающую роль в возникновении и развитии АС, а активные формы кислорода (ROS) и продукты их реакции являются важными медиаторами в сосудистой патологии и окислительном стрессе, в частности, супероксидные анионы [22].
22.Priestley JRC, Buelow MW, McEwen ST, Weinberg BD, Delaney M, Balus SF, Hoeppner C, et al. Reduced angiotensin II levels cause generalized vascular dysfunction via oxidant stress in hamster cheek pouch arterioles. Microvascular Res. . 2013;89:134–145. doi: 10.1016/j.mvr.2013.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
После того, как мышей ApoE –/– кормили диетой с высоким содержанием метионина, уровни супероксидных анионов увеличились (Рисунок 1F).
Зоны атерогенных поражений положительно коррелировали с уровнями супероксидных анионов (r=0,4428; P=0,018; Рисунок 1G), что указывает на то, что супероксидные анионы могут усиливать развитие атерогенных поражений.
Экспрессия EC-SOD подавляется Hcy in vivo и in vitro Сообщалось, что EC-SOD представляет собой важный защитный механизм от супероксидных анионов в артериальной микросреде [23]. Чтобы подтвердить влияние Hcy на экспрессию EC-SOD in vivo и in vitro, экспрессия EC-SOD была обнаружена в аорте и макрофагах.
1 результат только метионин
Наши результаты показали, что уровни мРНК EC-SOD были снижены у мышей ApoE –/–, которых кормили пищей с высоким содержанием метионина, и были повышены, когда мышей ApoE –/– кормили фолатом и витамином B 12 (Рисунок 2A).
2 результат = метионин + В9 и В12
Соответственно, уровни белка EC-SOD в аорте были снижены, когда мышей ApoE –/– кормили пищей с высоким содержанием метионина, и увеличились после того, как мышей кормили фолатом и витамином B 12 (Рисунок 2B).
В соответствии с экспрессией EC-SOD in vivo, уровни экспрессии EC-SOD были подавлены в макрофагах после обработки клеток различными концентрациями Hcy (Рисунок 2C,D).
Эти результаты предоставили достаточно доказательств того, что EC-SOD может быть связана с реакцией макрофагов на окислительный стресс, а фолат В9 и витамин B 12 могут защищать клетки от окислительного повреждения, вызванного токсичностью Hcy.
(A) После 15 недель лечения уровень мРНК EC-SOD в аорте мышей ApoE ‒/‒ определяли с помощью количественной ОТ-ПЦР.
(B) Анализ экспрессии EC-SOD в аорте мышей ApoE ‒/‒ методом вестерн-блоттинга.
(C) qRT-PCR использовали для измерения экспрессии уровней мРНК EC-SOD в макрофагах после обработки клеток различными концентрациями Hcy в течение 24 ч.
(D) Экспрессию белка EC-SOD измеряли с помощью анализа вестерн-блоттинга. * P < 0,05 и ** P < 0,001 по сравнению с контрольной группой; ▽ P < 0,01 по сравнению с группой A-control; △ P < 0,01 по сравнению с группой Meth в (A, B). △ P < 0,05 по сравнению с группой 100 мкМ Hcy в (C, D).
Антагон: 100 мкМ Hcy и 30 мкМ витамина B12 и 30 мкМ фолиевой кислоты.
Содержание супероксид-аниона измерялось после трансфекции рекомбинированным вектором и фрагментом интерференции РНК –596 для EC-SOD (рисунок 3F).
Содержание супероксид-аниона значительно увеличивалось в макрофагах после обработки Hcy. Напротив, содержание супероксид-аниона значительно уменьшалось при сверхэкспрессии EC-SOD и обработке Hcy (P<0,05). В совокупности эти результаты свидетельствуют о том, что EC-SOD играет важную роль в окислительном стрессе в макрофагах, вызванном Hcy. Гипометилирование EC-SOD в аорте мышей ApoE –/–, вызванное Hcy Активность транскрипции является ключом к определению уровней экспрессии генов в эукариотических организмах, которые подвержены различным факторам, таким как эпигенетическая модификация, включая метилирование ДНК [24]. Чтобы получить представление о лежащем в основе механизма EC-SOD в формировании атеросклеротической бляшки, мы трансфицировали несколько фрагментов 5′-фланкирующей области EC-SOD (–200/–1, –700/–1, –1100/–1, –1400/–1 и –2000/–1), которые были вставлены в вектор люциферазы светлячка pGL3. Анализ активности люциферазы показал, что область (–1100/–1), которая охватывает большую часть динуклеотида CpG промотора EC-SOD, имеет самую высокую активность промотора (рисунок 4A), что указывает на то, что регуляторные элементы для транскрипции EC-SOD, возможно, существуют в этой области.
В нашем предыдущем исследовании мы подтвердили, что DNMT1 играет важную роль в формировании атеросклеротической бляшки [25].
В настоящем исследовании мы обнаружили, что экспрессия мРНК DNMT1 была подавлена у мышей ApoE –/– (рисунок 4B).
Паттерн метилирования ДНК считается полезным молекулярным маркером АС из-за его роли в подавлении транскрипции ДНК [26].
Метилирование ДНК EC-SOD было проанализировано для изучения причины изменений экспрессии EC-SOD. Было обнаружено, что метилирование EC-SOD было подавлено в группе Meth и умерено после того, как мышей ApoE –/– кормили фолатом и витамином B 12 (рисунок 4C), что указывало на то, что Hcy подавлял метилирование ДНК EC-SOD.
Hcy suppresses EC-SOD methylation levels in the aortas of ApoE –/– mice
(A) Analysis of EC-SOD promoter activity. Different lengths of human EC-SOD 5′-flanking sequences were fused to the luciferase gene in a pGL-3.1. The fragments of promoter activity were subsequently analyzed by calculating the ratio of luminescence from the experimental reporter vector to the luminescence from a control vector (phRL-TK).
(B) The expression of DNMT1 mRNA in the aorta of mice was detected by qRT-PCR.
(C) The methylation status of EC-SOD in the aortas of mice was analyzed by methylation special touch down polymerase chain reaction.
(D) The concentrations of SAH and SAM in the aorta of mice were measured by HLPC.
(E) The ratio of SAM to SAH. * P<0.05 and ** P<0.01 vs the N-control group or the pGL3-basic group; △ P<0.05 vs the A-control group; ▽ P<0.05 vs the Meth group. pGL-3.1, pGL3-basic vector; M, methylated polymerase chain reaction band; U, unmethylated polymerase chain reaction band; SAM, S-adenosylmethionine; SAH, S-adenosylhomocysteine.