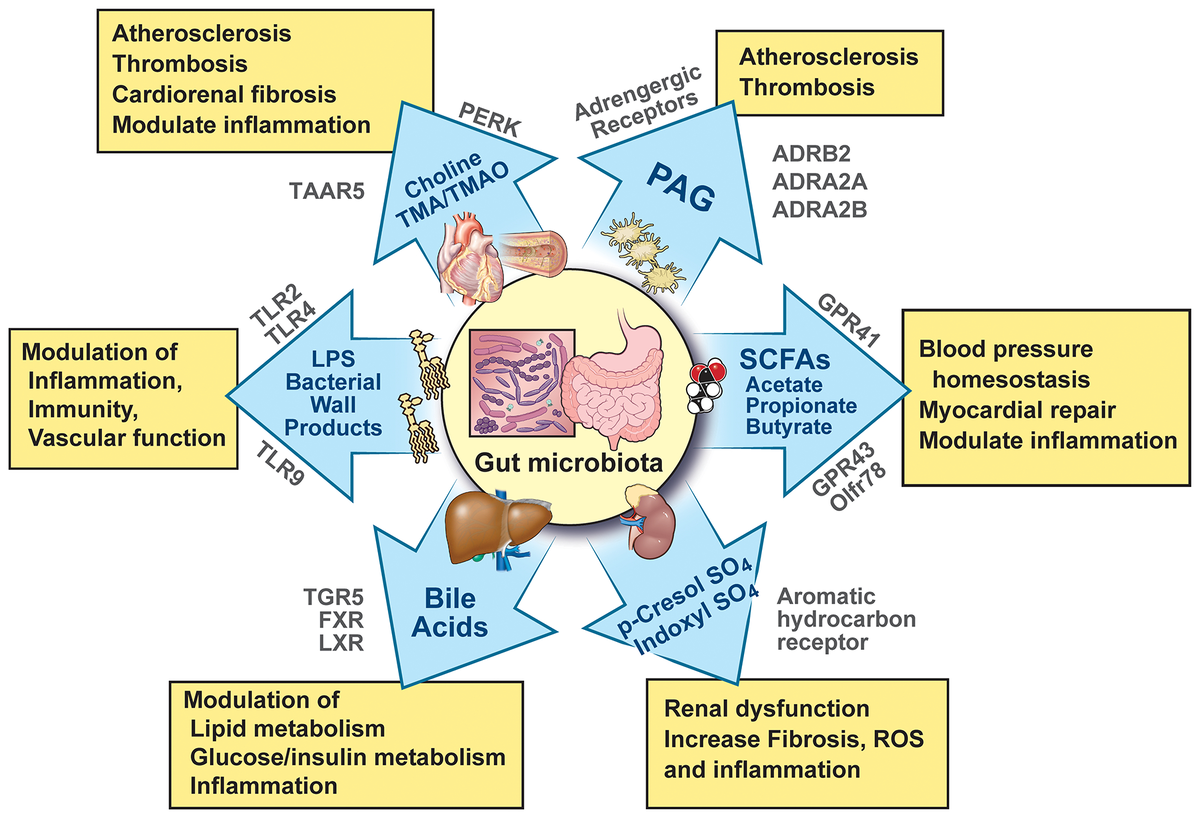
Молекулярные пути и принимающие рецепторы, которые связывают продукты и метаболиты, полученные от микробиоты кишечника, с фенотипами сердечно-сосудистых и кардиометаболических заболеваний. ADRA, адренэргический рецептор альфа; ADRB, адренэргический рецептор бета-бета; GPR, G-protein-coupled receptor; FXR, farnesol X receptor; LPS, lipopolysaccharide; LXR, liver X receptor; Olfr, olfactory receptor; PAG, phenylacetylglutamine; PERK, protein kinase, endoplasmic reticulum kinase; ROS, реактивные виды кислорода; SCFA, короткоцепная жирная кислота; TGR, рецептор со связью с белком Takeda G; TLR, рецептор с тибром; TMA, триметиламин; TMAO, триметиламин N-оксид; и TAAR, прибор с микроаминами.
Изменения состава кишечного бактериального сообщества и связь с сердечно-сосудистыми заболеваниями Благодаря достижениям в области технологий независимого от культуры секвенирования и биоинформатики подавляющее большинство исследований, связанных с кишечным микробиомом, оценивают изменения состава кишечного (часто фекального) микробного сообщества, связанные с различными болезненными состояниями.
Это привело к появлению большого количества ассоциативных данных, которые, хотя и полезны, ограничены с точки зрения исследования причинно-следственной связи. Хотя трудно определить действительно патогенное бактериальное сообщество, термин дисбактериоз использовался для описания дисбаланса состава кишечной микробиоты в пределах болезненного состояния или фенотипа. Многие исследователи сообщали о связи между фенотипами сердечно-сосудистых заболеваний и изменениями в относительном обилии определенных микробных таксонов или богатстве или разнообразии кишечных бактерий. Например, в ранних исследованиях бактериальная ДНК была обнаружена в атеросклеротических бляшках с сигнатурами, которые соответствуют таксонам, связанным с болезненными состояниями6, а изменения микробного состава были зарегистрированы у пациентов с многочисленными факторами риска сердечно-сосудистых заболеваний, включая гипертонию, дислипидемию, резистентность к инсулину и другие метаболические фенотипы10, 11. Изменения в составе микробиоты, ее разнообразии и богатстве являются ассоциативными, что затрудняет определение того, являются ли микробные изменения движущей силой болезненных состояний или, скорее, вызываются ими. Более того, обычно используемые методы секвенирования часто не достигают разрешения на уровне видов или штаммов, а анализы часто исключают менее распространенные микробы.
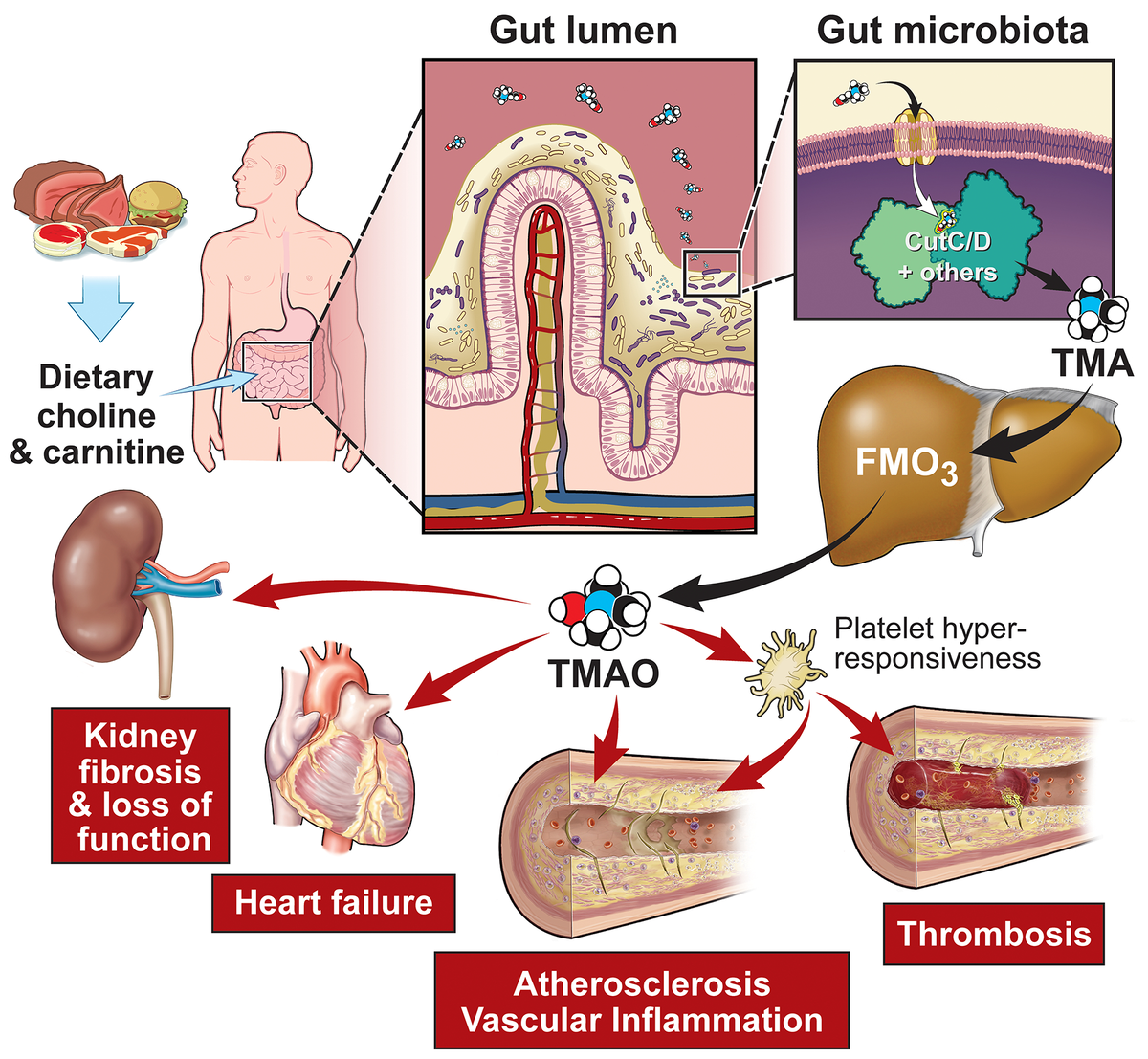
Исследования, сосредоточенные в первую очередь на распространенности, могут, таким образом, игнорировать микробные пути приобретения функции, которые способствуют заболеванию, несмотря на то, что они возникают из микробов, которые могут представлять лишь небольшую часть микробного сообщества. Недавний наглядный пример этого показан в серии исследований микробной трансплантации, разработанных для демонстрации прямой роли микробного cutC кишечника (основного микробного гена, ответственного за трансформацию холина → ТМА12, 13) в потенциале тромбоза хозяина14 (рисунок 2).
В этих исследованиях использовались мыши без микробов (GF), колонизированные синтетическим полимикробным сообществом, лишенным функциональной способности холина → ТМА, и добавление человеческого комменсала (C. sporogenes), генетически сконструированного так, чтобы либо обладать, либо не обладать функциональным микробным геном cutC. Хотя он присутствовал только в очень низкой численности (всего ~ 0,1%) в микробном сообществе толстой кишки, мутант C. sporogenes с усилением функции повысил уровни циркулирующего ТМА и ТМАО в организме хозяина, а также увеличил скорость образования тромбов и сократил время до прекращения кровотока после артериальной травмы in vivo14.
Проницаемость кишечника как потенциальный портал для продуктов, полученных из микробиоты кишечника, и воспаления
В здоровом состоянии барьерная функция кишечника поддерживается физическими факторами, включая плотные соединения между эпителиальными клетками, выработку слизи и иммунитет слизистой оболочки.
А у пациентов с сердечной недостаточностью мы часто наблюдаем отек стенки кишечника и нарушение барьерной функции18, 19.
- 18.Polsinelli VB, Sinha A, Shah SJ. Visceral congestion in heart failure: Right ventricular dysfunction, splanchnic hemodynamics, and the intestinal microenvironment. Curr Heart Fail Rep. 2017;14:519–528 [DOI] [PMC free article] [PubMed] [Google Scholar]
Существует несколько потенциальных методов лечения, которые могли бы улучшить патофизиологические изменения, относящиеся к правой оси сердца-кишечника при сердечной недостаточности.
Показаны потенциальные методы лечения, которые могут улучшить правый желудочек, легкие сосуды, кишечную микросреду и почку. Эти методы лечения являются теоретическими по большей части, и все еще необходимо тщательно изучить и доказать эффективность.
RVAD = устройство для оказания помощи правому вентральному; ERA = антагонисты рецепторов эндотелина; PDE5 = фосфодиэстерезия-5; sGC = растворимый гуанилат циклалы; FGF23 = фактор роста фибробластов-23; NHE3 = натрий-водорнеобменник З
- 19.Tang WHW, Li DY, Hazen SL. Dietary metabolism, the gut microbiome, and heart failure. Nat Rev Cardiol. 2019;16:137–154 [DOI] [PMC free article] [PubMed] [Google Scholar]
Низкий сердечный выброс при сердечной недостаточности приводит к ишемии кишечника, отеку и воспалению, что приводит к «протекающему» кишечному барьеру.
Это позволяет повышенному прохождению воспалительных бактериальных продуктов попадать в кровоток, вызывая хроническое слабовыраженное воспаление. Кроме того, это изменяет кишечную среду и влияет как на нормальное микробное сообщество, которое находится в кишечнике, так и впоследствии на продукты метаболизма этих бактерий.
Метаболические пути включают ферментацию неперевариваемой клетчатки в короткоцепочечные жирные кислоты, которые обладают защитными свойствами, уменьшая воспаление и улучшая сосудистый тонус.
Пищевые источники, включая холин, фосфатидилхолин, L-карнитин и другие содержащие метиламин питательные вещества, обеспечивают субстраты для опосредованного микробиотой производства триметиламина (ТМА).
Затем ТМА попадает в портальный кровоток и преобразуется семейством ферментов печеночного хозяина флавинсодержащей монооксигеназы (FMO) в триметиламин-n-оксид (ТМАО).
- ТМАО может способствовать развитию атерогенеза, тромбоза, заболеваний почек и сердечной недостаточности.
Кроме того, бактериальная трансформация желчных кислот может привести к изменению профилей желчных кислот, которые затем могут повлиять на системные воспалительные и фиброзные процессы. В совокупности эти процессы могут влиять на индивидуальную восприимчивость, тяжесть сердечной недостаточности.
Следуя концепции, нарушение барьерной функции кишечника приводит к транслокации бактериальных продуктов в кровообращение хозяина, что может привести к провоспалительному состоянию.
Многочисленные исследования показывают, что у пациентов с сердечной недостаточностью наблюдаются изменения в целостности кишечника, и что повышенный уровень провоспалительных цитокинов в крови коррелирует с тяжестью симптомов и более плохими результатами.
При наличии сердечной недостаточности перегрузка венозной жидкостью, адаптивная симпатическая активация, приводящая к перераспределению системного кровообращения, и низкий сердечный выброс способствуют отеку стенки кишечника и снижению перфузии слизистой оболочки.
Кишечная гипоперфузия при сердечной недостаточности изменяет функцию слизистой оболочки, о чем свидетельствует повышенная парацеллюлярная проницаемость и усиленное образование кишечной бактериальной биопленки.
Когда кишечный барьер нарушен, липополисахарид (ЛПС), происходящий от грамотрицательных бактерий, может попасть в кровоток хозяина, где он в основном распознается толл-подобными рецепторами (TLR) на поверхности иммунных клеток.
При связывании бактериальных лигандов TLR-сигнализация вызывает высвобождение провоспалительных цитокинов, которые организуют провоспалительное состояние у хозяина. Повышенные уровни ЛПС и других продуктов бактериальной стенки, предположительно полученных из микробиоты кишечника, были механически связаны с модуляцией воспаления, иммунитета и сосудистой функции (рисунок 1).
У пациентов с декомпенсированной сердечной недостаточностью уровень эндотоксина в крови выше, чем у стабильных пациентов.
Транслокация ЛПС из кишечника в этой ситуации подтверждается более высокими концентрациями эндотоксина в печеночной вене по сравнению с кровью, взятой непосредственно из желудочков.
Обнаружение кишечной микробиоты и метаорганизменных метаболитов на повышенных уровнях у пациентов с сердечно-сосудистыми заболеваниями или риском возникновения неблагоприятных сердечно-сосудистых событий может частично отражать изменения барьерной функции хозяина (рисунок 1).
В недавнем наблюдательном исследовании циркулирующие концентрации ЛПС были предикторами MACE в когорте пациентов с фибрилляцией предсердий, что свидетельствует о том, что транслокация эндотоксина влияет на осложнения сердечно-сосудистых заболеваний.
Стоит отметить, что средиземноморская диета была отрицательно связана с эндотоксемией в этом исследовании, что свидетельствует об участии кишечной микробиоты в барьерной функции кишечника хозяина.
Действительно, в доклинических моделях зондовое введение живой Akkermansia muciniphila мышам, питавшимся западной диетой, снижало проницаемость кишечника и снижало уровни фекального и циркулирующего ЛПС, что было связано со снижением атеросклероза аорты независимо от метаболизма липидов.
Хотя механизм изменений барьерной функции, вызванных Akkermansia, необходимо изучить, недавнее исследование на людях, подтверждающее концепцию, показало, что введение пастеризованной (но не живой) Akkermansia muciniphila в течение 3 месяцев приводило к снижению уровня ЛПС в плазме у людей с ожирением и метаболическим синдромом.
Хотя вмешательство не изменило массу тела, авторы наблюдали улучшение чувствительности к инсулину и дислипидемии в группе, получавшей Akkermansia.
Интересно, что структурные различия подтипов ЛПС из разных видов кишечных микробов были связаны с измененным распознаванием TLR и их влиянием на врожденный иммунитет хозяина.
Например, зондовое введение грамотрицательных бактероидов (vulgatus и dorei), которые продуцируют пента- и тетраацилированный липид А — в отличие от гексаацилированного липида А кишечной палочки, снижает воспаление толстой кишки, эндотоксемию и атеросклероз у мышей ApoE−/−31.
Следует отметить, что факторы, выходящие за рамки кишечной проницаемости, способствуют развитию сердечно-сосудистых заболеваний.
Существует множество заболеваний, при которых наблюдается повышенная «кишечная проницаемость», но не все такие заболевания демонстрируют повышенную связь с риском сердечно-сосудистых заболеваний.
Например, в то время как наличие отека стенки кишечника, вызванного сердечной недостаточностью, было связано с эндотоксемией и прогрессированием сердечно-сосудистых заболеваний, дефекты кишечного барьера, вызванные колитом и воспалительными заболеваниями кишечника, классически не связаны с повышенным риском сердечно-сосудистых заболеваний.
Эти наблюдения указывают на более сложную связь между целостностью стенки кишечника, изменениями в микробных сообществах кишечника и связью между системным воспалением хозяина и измененной восприимчивостью к развитию сердечно-сосудистых заболеваний.
Мы предполагаем, что различные механизмы «проницаемости кишечника» (например, воспалительные, отек стенки кишечника и т. д.) могут по-разному влиять на микробиом кишечника и патофизиологические процессы, которые модулируют восприимчивость к развитию сердечно-сосудистых заболеваний.
Улучшенное понимание роли бактериальных провоспалительных факторов кишечника в запуске системных воспалительных каскадов может помочь обеспечить новые терапевтические стратегии для улучшения ухода и стратификации риска.
Downstream Effects of Gut Microbiota-generated Short-chain Fatty Acid (SCFA) in Cardiovascular System
Желчные кислоты
Желчные кислоты традиционно рассматриваются как эмульгаторы жиров и жирорастворимых витаминов, способствующие всасыванию в кишечнике. Первичные желчные кислоты, такие как холевая и хенодезоксихолевая кислота (ХДХК), синтезируются путем окисления холестерина в печени.
Эти первичные желчные кислоты обычно реабсорбируются (>95%)127. Однако микробиота в толстой кишке может далее метаболизировать любые непереработанные желчные кислоты для производства вторичных желчных кислот, таких как дезоксихолат (ДХК), литохолат (ЛХК), урсодезоксихолат (УДХК) и многочисленные (не менее 50) другие128,129. Почти весь пул желчных кислот в толстом кишечнике состоит из вторичных желчных кислот с концентрацией до 1000 мкМ. Эти вторичные желчные кислоты также попадают в энтерогепатическую циркуляцию, а оставшаяся экскреция происходит через фекалии.
Многочисленные доказательства связывают различные аспекты воспаления с повышенным риском сердечно-сосудистых заболеваний32, 33. Роль воспалительных путей в сердечно-сосудистых заболеваниях недавно была подтверждена клиническим исследованием CANTOS. Введение канакинумаба, антитела к интерлейкину (ИЛ)-1β, показало, что ингибирование пути ИЛ-1β снижает риск сердечно-сосудистых событий независимо от снижения уровня липидов34. Таким образом, лечение и профилактика сердечно-сосудистых заболеваний с помощью иммуномодулирующей терапии кажутся многообещающими. Но воздействие на воспалительные пути несет риск оппортунистических инфекций, что может ограничить его использование у пациентов с множественными сопутствующими заболеваниями, что часто наблюдается у пациентов с сердечно-сосудистыми заболеваниями. Выявление кишечной микробиоты, которая вызывает иммунные реакции хозяина, участвующие в патогенезе сердечно-сосудистых заболеваний, может предоставить терапевтический путь для улучшения фенотипов сердечно-сосудистых заболеваний, вызванных воспалением. Желчные кислоты в основном являются производными кишечной микробиоты и служат модуляторами метаболизма хозяина Основная функция, исторически приписываемая желчным кислотам (ЖК), заключается в содействии эмульгированию и адсорбции жирорастворимых пищевых питательных веществ. Однако ЖК состоят из разнообразного набора структурно-специфичных видов, концентрации которых различаются на много порядков. Более поздние исследования показали, что структурно-специфичные и отдельные ЖК также играют дополнительные роли, включая, помимо прочего, модуляцию липидного метаболизма хозяина, метаболизма глюкозы/инсулина и воспаления (рисунок 1).
Первоначально синтезированные из холестерина в печени хозяина, первичные ЖК, которые представляют собой лишь небольшую часть общего пула ЖК, затем секретируются в просвет кишечника (двенадцатиперстной кишки), где последующие модификации, зависящие от кишечной микробиоты, участвуют в образовании необычайно большого набора видов ЖК. Организм поддерживает большой пул гидрофобных ЖК посредством обратного захвата в подвздошной кишке и посредством отрицательной обратной связи. Эти механизмы отрицательной обратной связи запускаются активацией фарнезоидного х-рецептора (FXR) и холестерина 7α-гидроксилазы35 или экспрессией кишечных желчных транспортеров36. BAs модулируют микробный состав кишечника посредством мощных антимикробных свойств, иммунных реакций37 и FXR38. А обструкция желчи может привести к синдромам избыточного роста бактерий39. Микробиота кишечника модифицирует первичные BAs посредством гидролиза желчных солей и дегидроксилирования желчной кислоты 7α, что приводит к образованию вторичных BAs, многие из которых выполняют гормоноподобные функции. После системной адсорбции некоторые вторичные BAs, в свою очередь, влияют на физиологию хозяина посредством взаимодействия с различными ядерными рецепторами хозяина, включая FXR, печеночный рецептор X (LXR), рецептор прегнана X (PXR) и специфические рецепторы, сопряженные с G-белком, такие как TGR5 (рисунок 1)40–44.
Нарушения динамических взаимодействий между диетой, микробиотой кишечника и конкретными BA могут, таким образом, способствовать кардиометаболическим фенотипам и восприимчивости к заболеваниям. Например, измененные уровни BA в плазме связаны с резистентностью к инсулину при диабете 2 типа45, 46, а модуляция сигнализации BA может способствовать улучшению метаболизма во время некоторых видов противодиабетического лечения47. Таким образом, измерение системных уровней BA может помочь в оценке потенциального вклада микробиоты кишечника в кардиометаболические заболевания46. Лучшее понимание того, как нарушения в профилях желчных кислот связаны с будущим развитием болезненных состояний или ответами на терапию. Функциональные исследования для определения того, связаны ли механистически кандидаты на желчные кислоты с этими процессами, являются многообещающими областями будущих исследований. Короткоцепочечные жирные кислоты (SCFA) и артериальное давление у хозяина SCFA (жирные кислоты с 5 атомами углерода или менее) могут быть продуктами метаболизма хозяина (например, ацетат48). Однако они также вырабатываются в больших количествах микробиотой кишечника посредством анаэробной ферментации пищевых волокон49, 50. Наиболее распространенные SCFA включают ацетат, пропионат и бутират, которые связаны с изменениями в гомеостазе артериального давления хозяина, восстановлением миокарда и воспалением (рисунок 1). Идея о том, что циркулирующие SCFA в значительной степени вырабатываются кишечными комменсалами, подтверждается исследованиями, показывающими, что уровни свободных SCFA практически не определяются в плазме, полученной от животных, не содержащих микробов51. Помимо того, что они действуют как источник энергии для эпителиальных клеток толстой кишки (например, колоноцитов), SCFA всасываются в портальную кровь и участвуют в различных процессах хозяина, включая липидный обмен, гомеостаз глюкозы, воспаление кишечника и нейрогенез52.
Связь между SCFA и ожирением в раннем возрасте была показана Чо и др.5. Воздействие антибиотиков во время отлучения от груди изменило микробные сообщества кишечника с увеличением метаболической способности производить ацетат, пропионат и бутират у мышей C57BL/6J. Приток этих короткоцепочечных жирных кислот в печень привел к существенным изменениям в регуляции метаболизма липидов печени и ожирению5. Более того, воздействие антибиотиков в раннем возрасте также было связано с изменениями в разнообразии микробиоты53, lasti
ng эффекты на иммунитет хозяина54 и кардиометаболические заболевания, такие как диабет55, 56. Интересно, что перенос микробиоты, нарушенной антибиотиками, к следующему поколению мышей привел к потере микробного богатства и изменениям в экспрессии метагеномных генов и восприимчивости к колиту57. Первоначальные клинические интервенционные исследования сообщили, что потребление клетчатки связано со снижением артериального давления58, и поддерживают идею о том, что SCFA участвуют в регуляции артериального давления. Ранние механистические исследования Плужника и коллег подтвердили эту идею, продемонстрировав, что рецепторы SCFA, связанные с G-белком, обонятельный рецептор 78 (Olfr78) и рецептор G-белка 41 (Gpr41) участвуют в регуляции артериального давления хозяина59–61. Будучи экспрессированным в гладкомышечных клетках сосудов, а также в юкстагломерулярном аппарате, Olfr78 опосредует высвобождение ренина и изменения сосудистого сопротивления, способствуя гипертонии60. Напротив, Gpr41 экспрессируется в сосудистом эндотелии и способствует снижению артериального давления61. Введение пропионата в отсутствие Gpr41 имело тенденцию к повышению артериального давления, в то время как оно вызывало выраженное падение артериального давления у мышей Olfr78−/−, что предполагает дифференциальную функцию обоих рецепторов в регуляции артериального давления, зависящей от SCFA60. Интересно, что когда пул SCFA был истощен антибиотиками у мышей Olfr78−/−, артериальное давление повышалось, что еще раз подтверждает защитную роль микробной генерации SCFA для балансировки сигнализации Olfr78. Однако общие гипотензивные эффекты у животных дикого типа можно объяснить как снижением сердечного выброса, так и потерей сосудистого сопротивления, проявляемого SCFA62.
Хотя подавление микробиоты с помощью антимикробных препаратов (т. е. плохо усваиваемых антибиотиков; рисунок 3) может служить ценным инструментом для демонстрации участия микробиома кишечника в фенотипе хозяина, это нецелесообразный подход для терапевтического воздействия на микробиом кишечника с целью достижения желаемого долгосрочного результата у хозяина из-за отбора сообществ кишечных микробов с устойчивостью к антибиотикам.
Короткоцепочечные жирные кислоты (SCFA) и артериальное давление у хозяина SCFA (жирные кислоты с 5 атомами углерода или менее) могут быть продуктами метаболизма хозяина (например, ацетат48).
Однако они также вырабатываются в больших количествах микробиотой кишечника посредством анаэробной ферментации пищевых волокон49, 50. Наиболее распространенные SCFA включают ацетат, пропионат и бутират, которые связаны с изменениями в гомеостазе артериального давления хозяина, восстановлением миокарда и воспалением (рисунок 1)
. Идея о том, что циркулирующие SCFA в значительной степени вырабатываются кишечными комменсалами, подтверждается исследованиями, показывающими, что уровни свободных SCFA практически не определяются в плазме, полученной от животных, не содержащих микробов51.
Помимо того, что они действуют как источник энергии для эпителиальных клеток толстой кишки (например, колоноцитов), SCFA всасываются в портальную кровь и участвуют в различных процессах хозяина, включая липидный обмен, гомеостаз глюкозы, воспаление кишечника и нейрогенез52.
Связь между короткоцепочечными жирными кислотами и ожирением в раннем возрасте была продемонстрирована Чо и соавторами5. Воздействие антибиотиков во время отлучения от груди изменило микробные сообщества кишечника с увеличением метаболической способности производить ацетат, пропионат и бутират у мышей C57BL/6J.
Приток этих короткоцепочечных жирных кислот в печень привел к существенным изменениям в регуляции метаболизма липидов печени и фенотипу ожирения5.
Более того, воздействие антибиотиков в раннем возрасте также было связано с изменениями в разнообразии микробиоты, длительным воздействием на иммунитет хозяина и кардиометаболическими заболеваниями, такими как диабет.
Интересно, что перенос нарушенной антибиотиками микробиоты следующему поколению мышей привел к потере микробного богатства и изменениям в экспрессии метагеномных генов и восприимчивости к колиту57. Первоначальные клинические интервенционные исследования показали, что потребление клетчатки связано со снижением артериального давления58, и подтверждают идею о том, что короткоцепочечные жирные кислоты участвуют в регуляции артериального давления.
Ранние механистические исследования Плужника и коллег подтвердили эту идею, продемонстрировав, что рецепторы SCFA, связанные с G-белком, обонятельный рецептор 78 (Olfr78) и рецептор G-белка 41 (Gpr41) участвуют в регуляции артериального давления хозяина.
Будучи выраженным в клетках гладких мышц сосудов, а также в юкстагломерулярном аппарате, Olfr опосредует высвобождение ренина и изменения сосудистого сопротивления, способствуя гипертонии. Напротив, Gpr41 выражен в эндотелии сосудов и способствует снижению артериального давления
Введение пропионата в отсутствие Gpr41 имело тенденцию к повышению артериального давления, в то время как оно вызывало выраженное падение артериального давления у мышей Olfr78−/−, что предполагает дифференциальную функцию обоих рецепторов в регуляции артериального давления, зависящей от SCFA60.
Интересно, что когда пул SCFA был истощен антибиотиками у мышей Olfr78−/−, артериальное давление повысилось, что еще раз подтверждает защитную роль микробной генерации SCFA для балансировки сигнализации Olfr78. Однако общие гипотензивные эффекты у животных дикого типа могут быть объяснены как снижением сердечного выброса, так и потерей сосудистого сопротивления, проявляемого SCFAs62.
Хотя подавление микробиоты антимикробными препаратами (т. е. плохо всасывающимися антибиотиками; Рисунок 3) может служить ценным инструментом для демонстрации участия микробиома кишечника в фенотипе хозяина, это не жизнеспособный подход для терапевтического воздействия на микробиом кишечника с целью достижения желаемого долгосрочного результата у хозяина из-за отбора сообществ кишечных микробов с устойчивостью к антибиотикам.
Многие дополнительные исследования подтвердили роль генерации короткоцепочечных жирных кислот кишечной микробиотой в модуляции артериального давления у хозяина. Например, перенос фекалий от гипертензивных (по сравнению с нормотензивными) доноров мышам без микробов выявил передачу повышенного артериального давления.
Но в другом исследовании трансплантация фекалий от нормотензивных солеустойчивых крыс Dahl гипертензивным солечувствительным крысам Dahl на самом деле усугубила гипертонию у реципиентов, что предполагает, что дополнительные генетические переменные хозяина могут взаимодействовать с микробными факторами для модуляции контроля артериального давления64. Также была предложена роль короткоцепочечных жирных кислот в гипертензивном повреждении конечных органов у мышей, которым вводили ангиотензин II.
Таким образом, многочисленные линии доказательств показывают, что микробное сообщество кишечника может влиять на регуляцию артериального давления у хозяина, и что короткоцепочечные жирные кислоты представляют собой по крайней мере один из микробных медиаторов, которые способствуют вазомоторному тонусу и артериальному давлению. Недавние исследования предоставили дополнительные доказательства того, что SCFAs участвуют в других процессах сердечно-сосудистых заболеваний, таких как ишемическое реперфузионное повреждение, восстановление сердца после инфаркта миокарда и нарушение артериальной податливости.
SCFAs представляют собой считывание метаболизма сахаридов всем микробным сообществом и часто служат конечными продуктами (поли)микробных катаболических путей. Поэтому их уровни могут отражать конвергенцию множественных микробных участников и конкурирующих путей. Дальнейшее понимание факторов, которые способствуют отдельным молекулярным видам SCFAs и рецепторов хозяина, воспринимающих их, является областью будущих исследований.
Особый интерес представляют недавние исследования, использующие нецелевую метаболомику, предполагающие, что дополнительные метаболиты плазмы, полученные из кишечной микробиоты, помимо SCFAs, также могут способствовать регуляции артериального давления хозяина.
В ходе этих исследований было обнаружено несколько структурно-специфических соединений, включая некоторые «уремические токсины», которые, как ранее сообщалось, активируют ренин-ангиотензин-альдостероновую систему (РААС) и были связаны с повышенным повреждением почек в модельных системах.
Считается, что все большее число продуктов, зависящих от микробиоты, включая уремические токсины, такие как п-крезолсульфат, индоксилсульфат и различные метаболиты ароматических аминокислот, потенциально изменяют метаболизм хозяина через специфические рецепторы, включая ароматический углеводородный рецептор (рисунок 3)
.Триметиламин N-оксид — метаболит метаорганизма, причинно связанный с сердечно-сосудистыми заболеваниями и метаболическими заболеваниями Почти десять лет назад Ван и др. впервые причинно связали кишечную микробиоту с пероральным приемом прекурсоров питательных веществ, выработкой ТМАО и риском сердечно-сосудистых заболеваний (рисунки 1 и 2)
Комбинация нецелевых метаболомных и механистических исследований на животных моделях использовалась для выявления малых молекул, уровни которых в крови связаны с риском сердечно-сосудистых заболеваний у людей и влияют на фенотипы, соответствующие сердечно-сосудистым заболеваниям, в животных моделях.
Было выявлено несколько аналитов, связанных с метаболизмом фосфатидилхолина, включая ТМАО, и было показано, что ТМАО как предсказывает риски сердечно-сосудистых заболеваний в нескольких клинических когортах, так и способствует ускоренному атеросклерозу (как и предшественники питательных веществ с неповрежденным кишечным микробиомом хозяина) в животных моделях.
Благодаря этим и другим исследованиям было показано, что образование ТМАО у людей и мышей происходит через многоступенчатый метаорганизменный путь, начинающийся с пищевых предшественников холина9, фосфатидилхолина8, 9 и карнитина7.
Они наиболее распространены в продуктах, встречающихся в западной диете, включая красное мясо, яичные желтки и другие продукты животного происхождения (рисунок 2).
Примечательно, что уровни в плазме или сыворотке каждого дополнительного предшественника питательных веществ ТМА, идентифицированного (т. е. продемонстрировавшего образование ТМА и ТМАО у хозяев через зависимый от микробиоты кишечника способ), включая бетаин, γ-бутиробетаин и триметиллизин (ТМЛ), все аналогично показали связь с рисками возникновения сердечно-сосудистых заболеваний в крупномасштабных клинических исследованиях, и эти связи, по-видимому, опосредованы ТМАО (поскольку их клиническая прогностическая ценность ослабевает с ТМАО в модели).
Кишечный микробный метаболизм предшественников питательных веществ, содержащих ТМА, начинается со специфических микробных лиаз ТМА, которые производят ТМА, пахучий газ, в качестве продукта.
Считается, что основная микробная холиновая ТМА-лиаза кодируется микробными генами cutC/D (гены кластера генов утилизации холина C (каталитические) и D)12.
Затем полученный ТМА транспортируется в печень через воротную вену и легко метаболизируется печеночными флавинмонооксигеназами хозяина (FMO) (в основном FMO376) в ТМАО9.
В кровотоке уровни ТМА обычно незначительны. Когда радиоактивно меченые ТМА или ТМАО перорально вводились добровольцам, 95% дозы выводилось через почки, причем большая часть представляла собой ТМАО. Было показано, что ТМАО усиливает атеросклероз в большинстве, но не во всех, механистических и модельных исследованиях на животных
Также было показано, что он способствует реактивности тромбоцитов и тромбообразованию, воспалению сосудов и активации инфламмасом, усилению сердечной недостаточности и фенотипам, связанным с ХБП92–95, в моделях на животных (рисунок 2).
У мышей E−/− диетическое дополнение холином привело к увеличению нагрузки атеросклеротическими поражениями, более высокой экспрессии рецепторов-мусорщиков (CD36 и рецептора-мусорщика A) в аорте, увеличению образования пенистых клеток макрофагов, нагруженных холестерином, и нарушению обратного транспорта холестерина in vivo.
Кроме того, ТМАО подавлял размер пула желчных кислот и, следовательно, клиренс холестерина у хозяина7. В соответствии с этими результатами и в дополнение к ним было показано, что снижение FMO3 ухудшает трансформацию ТМА в ТМАО, тем самым снижая уровни ТМАО в плазме и, одновременно, восстанавливая баланс холестерина.
Ранние исследования микробной трансплантации микробных сообществ слепой кишки от высокопродуцирующих ТМА инбредных мышей C57BL/6J реципиентам NZW/LacJ, устойчивым к атеросклерозу, продемонстрировали трансмиссивность вызванной холином диеты генерации ТМА и ТМАО, а также атеросклероза.
Однако не все исследования кормления предшественниками ТМАО показали схожие результаты, подтверждающие идею о том, что различия в микробных сообществах, присутствующих в хозяине, могут влиять на конечные наблюдаемые фенотипы.
Поразительная связь между циркулирующими уровнями ТМАО у субъектов и рисками тромботических событий, такими как сердечный приступ и инсульт, была засвидетельствована в многочисленных крупномасштабных клинических когортах
Это побудило механистические исследования как на людях, так и на мышах для изучения роли ТМАО в тромбозе. Чжу и др. обнаружили, что ТМАО изменяет сигнализацию кальция в тромбоцитах человека, повышая их чувствительность к субмаксимальной стимуляции агонистами (например, тромбином, АДФ, коллагеном).
Следовательно, повышенный тромботический потенциал наблюдался как в цельной крови, так и в моделях артериальных повреждений in vivo.
В последующем исследовании с кормлением людей здоровые добровольцы (как всеядные, так и веганы/вегетарианцы), которым перорально вводили холин, показали более высокий уровень ТМАО и сопутствующую повышенную реакцию тромбоцитов и агрегацию.
Важно, что более высокие уровни ТМАО были дозозависимо связаны с повышенной реакцией агрегации тромбоцитов, даже у субъектов, принимавших низкие дозы аспирина.
Это говорит о том, что у субъектов с повышенным уровнем ТМАО антитромбоцитарные эффекты аспирина могут быть ослаблены, что подчеркивает возможное участие ТМАО в реактивности тромбоцитов во время лечения и так называемой «резистентности к аспирину».
Механистическое участие метаорганизменного пути TMAO в функции тромбоцитов и потенциале тромбоза in vivo также изучалось с помощью генетических манипуляций с усилением и потерей функции, в том числе с геном хозяина FMO3 (рисунок 2). Многочисленные исследования подтвердили с помощью как генетического усиления (как глобальный трансген FMO3), так и потери (через антисмысловой олигонуклеотид к FMO3 и через глобальный нокаут FMO3) исследований функции на мышах, что манипуляция уровнями TMA и TMAO in vivo изменяет чувствительность тромбоцитов, скорость образования сгустка и потенциал тромбоза.
Более того, эксперименты по трансплантации микробов слепой кишки подтвердили, что протромботический фенотип, опосредованный богатой холином диетой, является передающимся признаком.
Ранние исследования Cracium et al. впервые идентифицировали кластер генов утилизации холина (cut) у человеческих комменсалов, кодирующих каталитические и регуляторные продукты генов CutC и CutD12.
Наличие микробных генов cutC/D в микробиоте человека связано со способностью генерировать ТМА из холина и с последующим накоплением ТМАО.
Важно отметить, что исследования с использованием стерильных мышей, колонизированных синтетическими микробными сообществами +/- генетически сконструированный мутантный человеческий комменсал cutC с усилением или потерей функции, подтвердили, что функционального микробного гена cutC достаточно для передачи генерации ТМА и ТМАО, а также потенциала тромбоза in vivo.
Таким образом, микробный ген cutC может представлять собой терапевтическую цель для предотвращения тромбоэмболических осложнений.
Недавние исследования показали, что помимо воздействия на функцию тромбоцитов, ТМАО вызывает экспрессию тканевого фактора (ТФ), инициатора внешнего свертывания, в эндотелиальных клетках in vitro.
Сосудистый ТФ способствует тромбозу и воспалению сосудов, особенно у пациентов с диабетом 2 типа, у которых более высокие уровни циркулирующего ТМАО.
Исследования на животных моделях по-прежнему необходимы для подтверждения вклада кишечной микробиоты и генерации ТМАО в изменения пути ТФ in vivo и в измененный потенциал тромбоза. Недавнее исследование показало, что отсутствие микробиоты было связано с уменьшением синтеза печеночного фактора Виллебранда (ФВ) и уменьшением роста тромба после повреждения сонной артерии у безмикробных мышей C57BL/6 по сравнению с традиционно выращенными маленькими матами.
Однако роль метаболитов метаорганизма, в частности ТМАО, в тромбозе, зависящем от ФВ, еще предстоит определить. Сосудистое воспаление критически вовлечено в патогенез атеросклероза и тромботических осложнений. Селдин и др. обнаружили, что острое введение физиологических уровней ТМАО мышам усиливает сосудистое воспаление (рисунок 2), что подтверждается
Активация эндотелиальных клеток аорты (восстановленная лазерной микродиссекцией), включая активацию сигнализации митоген-активируемой протеинкиназы (МАРК) и ядерной транслокации ядерного фактора (NF) kB, что приводит к последующей провоспалительной экспрессии генов.
Дополнительные результаты (стимулированное ТМАО сосудистое воспаление; Рисунок 2) наблюдались in vitro с использованием первичных эндотелиальных клеток аорты человека и гладкомышечных клеток сосудов.
После острой инъекции аорты мышей также показали повышенную экспрессию молекул сосудистой адгезии, таких как E-селектин или молекула межклеточной клеточной адгезии (ICAM)-1, даже когда ТМАО был выведен из кровообращения, что подразумевает устойчивое сосудистое воспаление. Кроме того, сообщалось, что ТМАО увеличивает окислительный стресс и сосудистое старение, которое характеризуется нарушением пролиферации и миграции клеток в эндотелиальных клетках пупочной вены человека.
В других исследованиях на животных моделях подавление микробиоты кишечника с помощью пероральных плохо всасывающихся антибиотиков было связано со снижением уровня ТМАО, улучшением эндотелиальной функции, снижением артериальной жесткости и снижением окислительного стресса.
Недавние исследования показали, что ТМАО может влиять на воспаление посредством праймирования и активации инфламмасомы NLRP3 в эндотелиальных клетках, а также артериальной сосудистой стенки у мышей, включая продукцию митохондриальных активных форм кислорода (ROS), тиоредоксина, взаимодействующего с белком (TXNIP) и лизосомальную дестабилизацию.
Однако точные механизмы, с помощью которых ТМАО вызывает активность инфламмасомы, еще предстоит изучить. Рецептор ТМА был идентифицирован давно как высокочувствительный обонятельный рецептор, называемый рецептором 5, ассоциированным со следовыми аминами (TAAR5)132. TAAR5 проявляет высокую специфичность к ТМА и не распознает ТМАО. В то время как ТМА сам по себе может способствовать провоспалительной сигнализации в сосудистой системе86, роль TAAR5 в сердечно-сосудистых заболеваниях требует дальнейшего изучения. Недавно было сообщено о рецепторе для ТМАО, который, как было показано, участвует в ТМАО-зависимых эффектах на метаболизм глюкозы и инсулина
Чен и др. показали, что ТМАО напрямую связывается с протеинкиназой R-подобной эндоплазматической ретикулум-киназой (PERK), основным компонентом развернутого белкового ответа – сигнального пути, который адаптирует клетку к стрессу ЭР133 (рисунок 1).
Пищевое дополнение ТМАО у животных дикого типа C57BL/6J индуцировало экспрессию PERK в печени, сопровождавшуюся повышением Fox01, ключевого фактора транскрипции в выделении инсулина и нарушением гликемического контроля.
Генетическая манипуляция продемонстрировала, что отсутствие печеночной PERK притупляло вызванное ТМАО повышение экспрессии Fox01 и улучшало гликемические индексы. Играет ли PERK роль в опосредованных TMAO эффектах на атеросклероз или тромбоз, остается неизвестным и является важной областью для дальнейшего изучения. Также интересно отметить, что стресс ER участвует в патогенезе многих фенотипов сердечно-сосудистых заболеваний. Участвуют ли TMAO и PERK в этих ассоциациях, также еще предстоит изучить.
- Теоретические преимущества TMAO Благодаря своему небольшому размеру и сочетанию гидрофильных и гидрофобных свойств TMAO ведет себя как хаотропный агент, способный изменять конформацию белка и потенциально служить аллостерическим модулятором белков.
Эти особенности могут иметь важные физиологические функции у хозяина (например, влиять на разворачивание белка или реакцию ER на стресс в клетках). У некоторых водных животных, включая подгруппу глубоководных рыб, ТМАО, как сообщается, действует как осмолит и защищает от дестабилизации белков, вызванной давлением.
Действительно, некоторые костистые рыбы могут использовать большие количества ТМАО для осморегуляции, а уровни ТМАО в плазме у рыб увеличиваются с глубиной среды обитания, достигая уровней до 400 ммоль/кг у лилий, пойманных на глубине 7000 м в южной части Тихого океана.
Было показано, что ТМАО стабилизирует белки при повышенном гидростатическом давлении и, как полагают, влияет на конформационные изменения белков, подобные тем, которые происходят при аллостерической регуляции, и модулирует эффекты внутриклеточного молекулярного скопления.
Таким образом, накопление ТМАО как у глубоководных существ, так и у млекопитающих может представлять собой адаптивный механизм для воздействия на стабильность белков и внутриклеточные сигнальные процессы. В других исследованиях сообщалось, что ТМАО участвует в адаптивной реакции избегания замораживания на экстремальный холод — другими словами, он действует как антифриз — например, у ньюфаундлендской корюшки.
Эти рыбы обладают способностью повышать осмолярность плазмы за счет сезонного накопления ТМАО и других органических растворенных веществ, что снижает точку замерзания жидкостей организма и позволяет им выживать при отрицательных температурах.
Недавно была выдвинута гипотеза, что стабилизация белка, вызванная ТМАО, может играть роль в защите кардиомиоцитов от колебаний гидростатического давления во время сердечной недостаточности.
Напротив, высокие концентрации ТМАО иногда могут также нарушать функцию, как сообщалось об активности актомиозина.
Хотя наши знания о том, как микробиота влияет на сердечно-сосудистые заболевания, все еще рудиментарны, скорость, с которой появляются новые открытия, впечатляет.
Как уже отмечалось, существуют неопровержимые доказательства того, что процессы, происходящие от микробиоты кишечника, в целом связаны с многочисленными фенотипами, связанными с сердечно-сосудистыми заболеваниями, включая, помимо прочего, атеросклероз, реактивность тромбоцитов и тромботический потенциал, артериальное давление, липидный обмен, ожирение, гомеостаз глюкозы и сосудистое воспаление.
Исследовательские подходы включали широкий спектр исследований трансплантации микробиоты, как животных, так и людей, диетические вмешательства в колонизированных состояниях по сравнению с подавленными антибиотиками (или безмикробные мыши), прямое предоставление (диета или через инфузию) определенных метаболитов микробиоты кишечника, а также как генетические, так и фармакологические исследования, нацеленные на множественные компоненты метаорганизменных путей (включая гены микробиоты кишечника, а также как ферменты трансформации хозяина, так и рецепторы конечных органов).
Новые терапевтические подходы, нацеленные на микробиоту кишечника для лечения и профилактики сердечно-сосудистых заболеваний, представляют собой захватывающие области исследований.
Разработка нелетальных микробных ингибиторов, нацеленных на конкретные пути, но при этом демонстрирующих ограниченное системное воздействие на хозяина, является лишь одним из новых и потенциально многообещающих терапевтических подходов.
Другие включают, но не ограничиваются диетическими вмешательствами, пробиотиками и/или пребиотиками, которые, как мы надеемся, могут быть использованы для «терраформирования» микробного сообщества с целью изменения его функционального выхода на пользу хозяину.
Как и в случае с любым терапевтическим средством, для проверки новых терапевтических средств, нацеленных на микробиом кишечника, потребуются крупные перспективные интервенционные исследования.