Недавно была также предложена роль матриксной меди в новом пути клеточной смерти, называемом купроптозом
Медь (Cu) необходима эукариотам для метаболизма кислорода, детоксикации радикалов кислорода и поглощения железа (Fe)
Генетические исследования четко установили, что экспорт является основной формой защиты от токсичности Cu, поскольку клетки, лишенные ATP7A, значительно более чувствительны к избытку Cu по сравнению с теми, у которых отсутствуют металлотионеины [16].
[16].Gudekar N, Shanbhag V, Wang Y, Ralle M, Weisman GA, Petris MJ, Metallothioneins regulate ATP7A trafficking and control cell viability during copper deficiency and excess, Scientific Reports 10(1) (2020) 7856. [DOI] [PMC free article] [PubMed] [Google Scholar]
Вывод: если в клетке есть металлопротеин, зависимый от Меди (СU) ,то эта клетка страдает из-за избытка/недостатка меди
Мембранно-связанные компартменты внутри клетки дополнительно способствуют ограничению цитозольной Cu и минимизации токсичности, выступая в качестве участков секвестрации для уменьшения повреждения липидов и белков и предотвращения ненадлежащего неправильного металлирования участков связывания кофакторов цитозольных ферментов.
Однако при этом органеллы принимают на себя риск повреждения, вызванного Cu.
Cu проникает в клетку через высокоаффинный плазматический мембранный транспортер CTR1 (Cell transporter), а затем распределяется по различным мишеням через металлошапероны (ATOX1, CCS) или альтернативные лиганды (CuL, GSH).
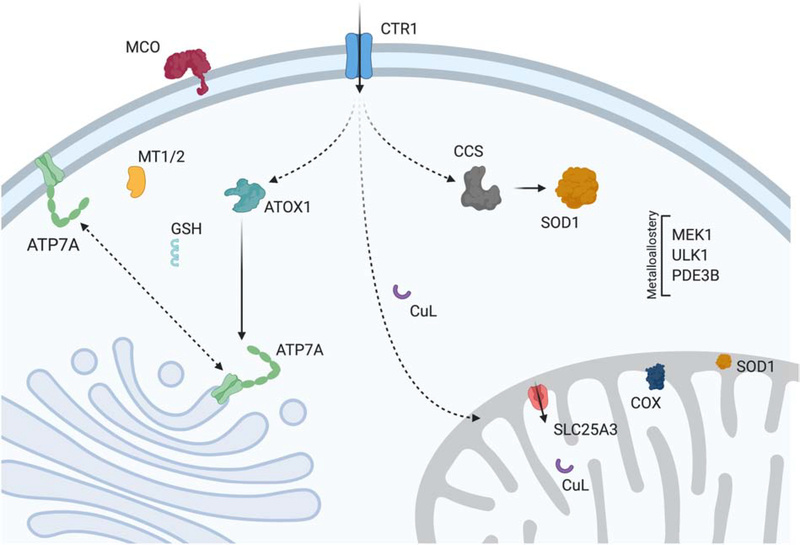
Cu проникает в транс-сеть Гольджи через взаимодействие между ATOX1 и ATP7A. После попадания Cu в цистерны он может быть включен в многочисленные ферменты, включая мульти-Cu оксидазы (MCO). В митохондриях Cu транспортируется в матрикс с помощью SLC25A3 для хранения.
После запуска неизвестным механизмом Cu транслоцируется обратно через IM к локализованным в IMS металлошаперонам, которые используют его для облегчения сборки цитохром с оксидазы (COX) и созревания супероксиддисмутазы (SOD1).
Cu в цитозоле используется в качестве кофактора SOD1 для защиты от окислительного стресса, а CCS требуется для металлирования этого сайта и образования необходимой дисульфидной связи.
Цистеин- аминокислота- важна, дает S-S группу для строительства супероксиддисмутазы, переноса меди и стабилизации мембран органелл
Когда Cu достигает избыточных уровней, он связывается металлотионеином (MT1/2), и ATP7A перемещается в PM(плазматическая мембрана), где он действует как экспортер Cu. В определенных типах клеток ATP7A заменяется на ATP7B, который перемещается в экзоцитарные везикулы во время стресса Cu, чтобы способствовать его выведению через желчь (показано ниже)
Динамические пулы Cu внутри клетки связываются в аллостерических сайтах, которые регулируют активность киназ MEK1 и ULK1, а также фосфодиэстеразы PD3EB в адипоцитах.
Механизмы, лежащие в основе этиологии заболевания и тканеспецифической экспрессии этих транспортеров, подробно рассматриваются в других работах [13, 14].
Митохондриальные ферменты IM и IMS собираются в результате согласованных действий нескольких металлошаперонов.
COX собирается в модули, которые впоследствии объединяются для получения зрелого голоферментного комплекса. COX1, который содержит сайт CuB и кофакторы гема, собирается как большой модульный комплекс, включающий SURF1, COA1 и COX11.
COX19 участвует в восстановлении Cu-связывающих цистеинов и активации COX11.
Сайт CuA собирается с помощью сложного набора окислительно-восстановительных взаимодействий, которые в конечном итоге позволяют SCO1 вставлять Cu в apo-COX2. COX20 физически взаимодействует с apo-COX2 при его вставке в IM, а COA6 и SCO2 работают согласованно, гарантируя, что цистеины активного сайта в SCO1 и COX2 поддерживаются в восстановленном состоянии во время созревания сайта CuA.
COX17 поставляет Cu, необходимую для сборки модулей COX1 и COX2. CMC1 также участвует в сборке COX, и генетические исследования на дрожжах подтверждают дополнительную роль CMC1 в продвижении активности SOD1.
CCS доставляет Cu в SOD1 и образует дисульфидную связь, необходимую для его активности. Предполагается, что Cu, используемая в этих реакциях, поступает из митохондриального матрикса.
Cu импортируется SLC25A3 после доставки через цитозоль неизвестным небелковым лигандом (CuL).
Альтернативная гипотеза заключается в том, что Cu хранится в комплексе IMS (Cu5000), который напрямую взаимодействует с металлошапероновыми белками, находящимися в этом отсеке.
Цистеин содержащий шапероны IMS
Митохондриальные ферменты IM и IMS собираются в результате согласованных действий нескольких металлошаперонов.
COX собирается в модули, которые впоследствии объединяются для получения зрелого голоферментного комплекса.
COX1, который содержит сайт CuB и кофакторы гема, собирается как большой модульный комплекс, включающий SURF1, COA1 и COX11.
COX19 участвует в восстановлении Cu-связывающих цистеинов и активации COX11.
Сайт CuA собирается с помощью сложного набора окислительно-восстановительных взаимодействий, которые в конечном итоге позволяют SCO1 вставлять Cu в apo-COX2. COX20 физически взаимодействует с apo-COX2 при его вставке в IM, а COA6 и SCO2 работают согласованно, гарантируя, что цистеины активного сайта в SCO1 и COX2 поддерживаются в восстановленном состоянии во время созревания сайта CuA.
COX17 поставляет Cu, необходимую для сборки модулей COX1 и COX2. CMC1 также участвует в сборке COX, и генетические исследования на дрожжах подтверждают дополнительную роль CMC1 в продвижении активности SOD1. CCS доставляет Cu в SOD1 и образует дисульфидную связь, необходимую для его активности.
Предполагается, что Cu, используемая в этих реакциях, поступает из митохондриального матрикса.
Cu импортируется SLC25A3 после доставки через цитозоль неизвестным небелковым лигандом (CuL).
Альтернативная гипотеза заключается в том, что Cu хранится в комплексе IMS (Cu5000), который напрямую взаимодействует с металлошапероновыми белками, находящимися в этом отсеке.
Цистеин, содержащий шапероны IMS (например, COA6, CMC1, COX17, COX19, CCS, SOD1), импортируется в IMS через комплекс TOM внешней мембраны и дисульфидный релейный аппарат MIA4/ERV1.
Cu необходим, но становится токсичным, когда его содержание превышает клеточную способность связывать Cu в инертных комплексах.
В митохондриях токсичность Cu приводит к повреждению липидов, окислению белков, окислительно-восстановительному дисбалансу из-за неправильного связывания с богатыми цистеином участками и истощению восстановленного глутатиона (GSH).
Кроме того, Cu разрушает и вытесняет Fe из открытого кластера FeS, содержащего ферменты.
Было показано, что Cu инактивирует ферредоксин и аконитазу, которые оба локализованы в матриксе.
Cu также инактивирует цитозольные ферменты, необходимые для синтеза лейцина (LEU1). В грибковых моделях делеция гомолога ABCB7 (Atm1), ответственного за транспортировку промежуточного продукта FeS неизвестной идентичности (FeS-X), увеличивает токсичность Cu и подчеркивает, что цитозольные мишени также являются важным аспектом токсичности Cu.
A) Неповрежденная 14-субъединичная структура человеческой цитохром с-оксидазы (PDB: 5z62), представленная в виде мультяшного остова с 3 митохондриально-кодируемыми субъединицами, выделенными синим цветом.
B) Структура COX с удаленными митохондриальными субъединицами для демонстрации гема и кофакторов Cu.
C) Увеличенное изображение двухъядерного участка CuA, гема a и смешанного металлогемового участка a3-CuB.
D) Мультяшное изображение половины димера SOD1 (PDB: 2c9v), показывающее его кофакторы Cu и цинка.
Открытие того, что ионофоры, такие как элескломол, могут использоваться для доставки Cu и восстановления функции в многочисленных животных и клеточных моделях расстройств, связанных с Cu, стало крупным достижением в этой области [28].
[28].Soma S, Latimer AJ, Chun H, Vicary AC, Timbalia SA, Boulet A, Rahn JJ, Chan SSL, Leary SC, Kim BE, Gitlin JD, Gohil VM, Elesclomol restores mitochondrial function in genetic models of copper deficiency, Proc Natl Acad Sci U S A 115(32) (2018) 8161–8166. [DOI] [PMC free article] [PubMed] [Google Scholar]
Но не рекомендую без клинического исслежования применять как "воду" в каждодневном рационе питания. Это требует консультаций со специалитсами....
Хотя ранее элескломол тестировался в гораздо более высоких концентрациях в качестве терапевтического средства для лечения рака, было обнаружено, что применение исключительно низких концентраций элескломола усиливает поглощение Cu и высвобождение внутренних запасов Cu, что позволяет устранить дефекты, связанные с Cu, в ряде субклеточных компартментов в нескольких моделях дрожжей и млекопитающих [28–31].
Кроме того, прямая инъекция связывающего Cu ионофора ATSM находится в фазе 2/3 клинического испытания (clincaltrials.gov: NCT04082832) для частичной коррекции симптомов заболевания у пациентов с боковым амиотрофическим склерозом (БАС) [32]. Однако все ионофоры и родственные им соединения токсичны при использовании в высоких концентрациях. У дрожжей такие соединения, как 2-(6-бензил-2-пиридил)хиназолин, вызывают значительное клеточное и митохондриальное накопление Cu, что приводит к повышенной токсичности [33]. Cu-elesclomol может вызывать гибель клеток в системах млекопитающих, и скрининг CRISPR по всему геному для супрессоров, устойчивых к этому ионофору, идентифицировал митохондриальный ферредоксин как цель, которая может модулировать токсичность [34].
Это открытие привело к описанию Cu-elesclomol-зависимой формы клеточной смерти, вызываемой ферредоксином, называемой купроптозом, которая происходит независимо от известных маркеров апоптоза и ферроптоза [34].
[34].Tsvetkov P, Detappe A, Cai K, Keys HR, Brune Z, Ying W, Thiru P, Reidy M, Kugener G, Rossen J, Kocak M, Kory N, Tsherniak A, Santagata S, Whitesell L, Ghobrial IM, Markley JL, Lindquist S, Golub TR, Mitochondrial metabolism promotes adaptation to proteotoxic stress, Nat Chem Biol 15(7) (2019) 681–689. [DOI] [PMC free article] [PubMed] [Google Scholar]
Резкое различие в результатах, от поразительного фенотипического спасения фатальных нарушений обработки Cu до потенцирования гибели клеток, подчеркивает, что регуляция Cu в митохондриях имеет решающее значение для нормальной клеточной физиологии, рисунок 3.
Поэтому этот обзор фокусируется на митохондриальном гомеостазе Cu и на том, как органелла влияет на другие пути обработки Cu в клетке.
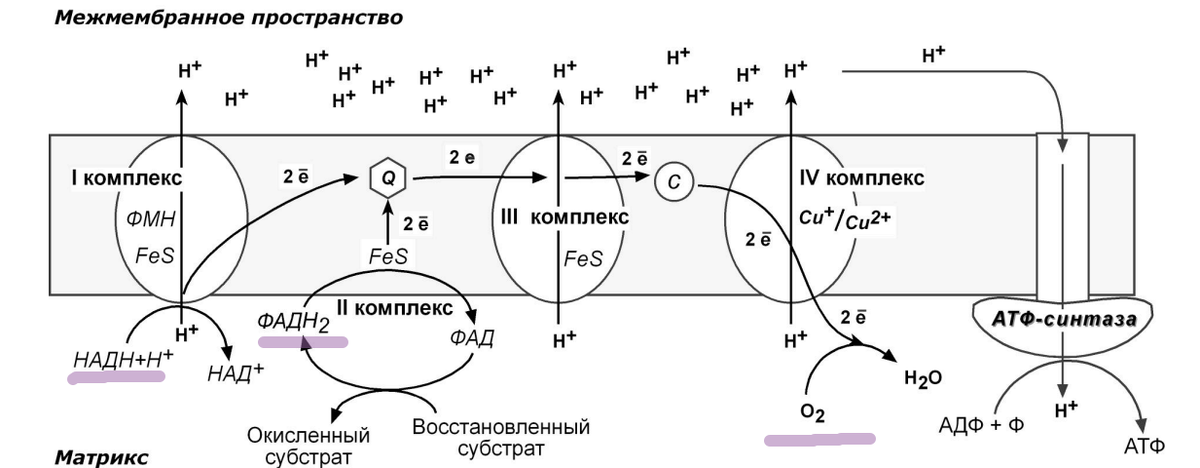
COX млекопитающих содержит 14 субъединиц, две из которых связывают 3 окислительно-восстановительных центра, необходимых для переноса электронов и перекачки протонов, рисунок 4.
Каталитическое ядро состоит из кодируемых митохондриями субъединиц COX1, COX2 и COX3.
Биогенез COX требует >25 дополнительных белков, известных как факторы сборки COX [35].
Различные факторы сборки COX
Трансляция COX1 и вставка в IM опосредованы набором специфических трансляционных активаторов и шаперонов, которые образуют комплекс, известный как MITRAC (промежуточный продукт сборки регуляции митохондриальной трансляции COX)
Супероксиддисмутаза: SOD1
SOD1 катализирует превращение радикала в молекулярном кислороде в перекись водорода (H2O2) посредством попеременного восстановления и окисления Cu, рисунок 4 [51].
Затем H2O2 независимо детоксифицируется каталазой и глутатионпероксидазой.
В то время как >90% от общего количества клеточного SOD1 находится в цитозоле, около 5% локализуется в IMS [52].
Митохондриальная локализация SOD1, по-видимому, зависит от механизма типа «fold-and-trap», поскольку для удержания белка при импорте в IMS требуется, чтобы SOD1 локализовался совместно с его родственным шапероном CCS и чтобы был доступен его кофактор Cu [51–53].
SOD1 не имеет традиционной митохондриальной последовательности нацеливания на своем аминоконце или внутреннего аминокислотного мотива, который нацеливает его на органеллу.
SOD1 должен быть свободен от металла и иметь восстановленную дисульфидную связь для перемещения через внешнюю мембрану (OM) и взаимодействия с релейной системой MIA40/ERV1.
Относительная приоритетность и распределение Cu в COX и SOD1 в IMS, по-видимому, регулируется, по крайней мере, частично фактором сборки COX CMC1, поскольку манипулирование уровнями экспрессии CMC1 влияет на количество ферментативно активного SOD1 [59, 60].
Однако механизмы, которые распределяют Cu в IMS и отдают ей приоритет для сборки COX или созревания SOD1, неизвестны.
Матрица Cu
Доставка Cu в митохондрии и накопление в них не зависит от присутствия и активности COX или SOD1 [61]. Общая концентрация Cu
Для доставки Cu в COX и IMS-SOD1 могут существовать два альтернативных пути.
Путь A изображает путь хранения матрицы, где Cu, который был привлечен в матрицу SLC25A3 для хранения, транспортируется обратно в IMS неопознанным транспортером для созревания фермента. Существование этого пути подтверждается многочисленными исследованиями в условиях стресса Cu.
Путь B изображает путь прямого доступа, где шапероны связывают Cu, когда он попадает в IMS, или получают к нему доступ после хранения в этом отсеке для поддержки созревания фермента. Хранение в IMS может быть облегчено комплексом Cu5000, который первоначально был выделен из митохондрий мозга.
Переход между этими двумя состояниями требует связывания субстрата, а относительная сила взаимодействий солевых мостиков определяет, действует ли транспортер как унипортер или как обменник. Сильные солевые мостики IMS и матрицы потребуют обмена субстрата для сброса в противоположное состояние. Любые более слабые солевые мостики позволят транспортеру быть однонаправленным.
A) Мультяшное представление остова Pic2, смоделированное на основе c-состояния обменника АДФ/АТФ (PDB:4C9G) B) Мультяшное представление остова Pic2, смоделированное на основе m-состояния обменника АДФ/АТФ (PDB:6GCI). C) Поверхностная визуализация модели Pic2 в модели c-состояния, смотрящей вниз по каналу со стороны IMS IM, подчеркивающей расположение остатков Cys21, Cys29, Cys44, His33, Met275, Arg175 и Lys183 желтым, зеленым и розовым цветами. D) Поверхностная визуализация модели Pic2 в модели m-состояния, подчеркивающей остатки, перечисленные в C).
Генетическая и фармакологическая манипуляция митохондриальной Cu в моделях млекопитающих приводит к резкому ремоделированию клеточного гомеостаза Cu.
Белки SCO генерируют окислительно-восстановительный сигнал, который частично передается в цитозоль посредством COX19, чтобы вызвать перемещение части пула ATP7A в PM для облегчения экспорта Cu.
Кроме того, сигнал(ы), опосредованный SCO, влияет на импорт Cu через CTR1, способствуя его протеасомной деградации или предотвращая его локализацию в PM. В то время как сочетание этих двух событий приводит к тому, что клетки-мутанты SCO становятся глубоко дефицитными по Cu, их митохондриальный пул Cu сохраняется. Напротив, удаление SLC25A3 вызывает дефект митохондриальной Cu, который запускает сигнал, который приводит к снижению уровней CCS и цитозольной активности SOD1. Мы предполагаем, что это повлияет на известное взаимодействие между SOD1 и казеинкиназой 1γ (CK1γ), тем самым регулируя утилизацию глюкозы в этой мутантной клеточной линии.
Основные моменты.
Митохондрии накапливают медь в своей матрице через белки семейства митохондриальных переносчиков Pic2/SLC25A3 для возможного созревания купроэнзимов цитохром с оксидазы и супероксиддисмутазы.
Регуляторные роли меди и купропротеинов, находящихся в митохондриях, продолжают расширяться за пределы органеллы, связываясь с модуляцией клеточного поглощения и экспорта меди и облегчением межорганной коммуникации.
Роль митохондриальных сигналов в регуляции содержания клеточной меди, предлагаемые механизмы транспорта меди в органеллу изучаются в контексте эволюционного происхождения путей гомеостаза меди.
Выводы и будущие направления
Существующие данные свидетельствуют о том, что митохондрии сохраняют выделенный пул Cu для возможного использования при сборке COX и SOD1 в IMS.
Матричное хранение предотвращает распределение в другие Cu-зависимые процессы, и важность поддержания этого пула могла привести к эволюции сигнального каскада, который делает митохондрии приоритетными для гомеостаза Cu.
Идентификация Pic2 и SLC25A3 как переносчиков Cu и фосфата с двойной функцией открывает много возможностей для исследования того, как клеточный гомеостаз Cu зависит от специфического истощения этого пула. Хелаторы являются ценным инструментом для исследования этих путей; однако они одновременно ограничивают несколько путей, и это может усложнить интерпретацию Cu-зависимых сигнальных каскадов. Поэтому разумно ожидать, что клеточные линии, истощенные по отдельным Cu-обрабатывающим белкам, таким как SLC25A3, будут более информативны в отношении открытия новых путей, регулируемых Cu.
Кроме того, путем идентификации дополнительных членов семейства MCF, которые транспортируют Cu, можно будет более подробно изучить, как ион металла перемещается между различными компартментами органелл. Остается несколько основных нерешенных вопросов относительно митохондриальной обработки Cu и ее влияния на гомеостаз Cu в здоровых и больных состояниях. Как Cu перемещается обратно в IMS?
Как регулируется этот экспорт? Можно предположить, что причина, по которой генетические скрининги не смогли идентифицировать дополнительные транспортеры Cu, заключается в том, что, возможно, Pic2/SLC25A3 может изменить направление транспорта с этим изменением направленности в зависимости от взаимодействующих партнеров.
В этой модели взаимодействия с шаперонами IMS могут регулировать высвобождение Cu обратно в IMS для сборки COX и SOD1.
Для решения этого вопроса требуется полный каталог белков, которые взаимодействуют с Pic2/SLC25A3. Однако эти усилия были оспорены тем фактом, что SLC25A3 является частым загрязнителем в сложных белковых смесях, полученных с использованием методов физического вытягивания [125].
Может ли транспорт фосфата и Cu быть функционально разделен в SLC25A3?
Как митохондриальный сигнал передается в цитозоль для скоординированной регуляции активности различных клеточных путей гомеостаза Cu?
В настоящее время представляется, что регуляция часто включает деградацию мишеней. Итак, как протеасома или другие протеазы получают сигнал для избирательной деградации определенных белков Cu?
Какие другие физиологические функции нарушаются при манипулировании митохондриальными пулами Cu?
Как регулируется токсичность Cu в митохондриальном матриксе относительно FeS, особенно в свете купроптоза?
Ответы на эти вопросы, а также расширение наших знаний о металлоаллостерии митохондриальных мишеней станут основными достижениями в области митохондриальной Cu в течение следующих нескольких лет.