МГЭ очень разнообразны: есть сложная иерархически организованная классификация. Во всей полноте мы вас ею, естественно, «грузить» не будем. Расскажем лишь о двух основных классах. Первый это ДНК-транспозоны.
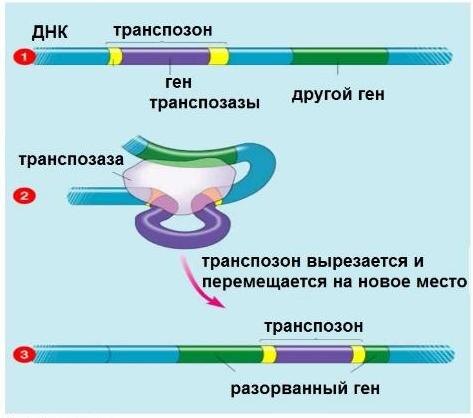
Они имеют небольшие размеры, ограничены с двух сторон короткими инвертированными повторами нуклеотидов (например, agtaaa ... tttact). Повторы могут «слипаться» друг с другом на основе принципа комплементарности. При этом транспозон приобретает вид петли, которая легко вырезается из молекулы ДНК соответствующим ферментом и переносится на новое место. Принцип перемещения ДНК-транспозонов по геному максимально простой: cut and paste (вырезаться-вставиться). Они буквально перепрыгивают с места на место. Именно их по-английски называют jamping genes (прыгающие гены).
Второй класс МГЭ – ретротранспозоны.
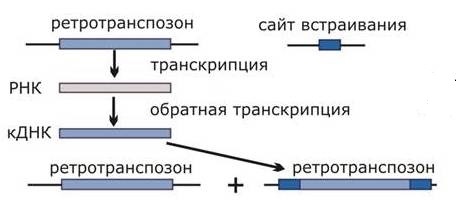
У них сразу два принципиальных отличия от ДНК-транспозонов. (1) Они не вырезаются из хромосомы, как это делают транспозоны. Принцип их действия – copy and paste (скопироваться и вставиться). Иными словами, они не только скачут по геному, но еще и плодятся. (2) Механизм их перемещения основан на существовании реакции обратной транскрипции - синтеза нити ДНК на РНК. Отсюда их название – РЕТРО. В процессе своего перемещения сначала синтезируют цепь РНК, с которой затем с помощью фермента обратной транскриптазы синтезируется комплементарная цепь ДНК. После удвоения двуцепочечная ДНК может встраиваться в различные районы хромосом хозяина. Ретротранспозоны значительно более разнообразны по размеру и структуре, чем ДНК-транспозоны. Повторим еще раз.
Есть ДНК-транспозоны, которые вырезаются и вставляются на новое место (cut and paste). При этом происходят две мутации, делеция (вырезка) и инсерция (вставка), но размер генома не увеличивается.
И есть ретроранспозоны, которые не вырезаются из ДНК, а лишь копируются и вставляются на новое место (copy and paste). При этом происходит лишь одна мутация, инсерция (вставка), с увеличением размера генома на размер вставки. Эта информация скоро понадобится нам, когда мы будем сравнивать геномы хвойных с геномами других растений.
Таким образом, мобильные генетические элементы сочетают в себе признаки эгоистических фрагментов ДНК («геномных паразитов») и важных составных частей генома, придающих ему пластичность и приспособляемость. Поэтому перед геномом хозяина стоит двуединая задача. С одной стороны, он обязан держать паразитов в узде, чтобы те не «хулиганили» и не слишком размножались. С другой стороны, он должен по возможности превратить их в своих «домашних животных», а затем использовать в этом качестве. Как эти задачи решены у хвойных в сравнении с их лиственными конкурентами? Этот вопрос изучен в уже использованной выше работе (Liu, El-Kassaby, 2019):
Среди лиственных деревьев их 30, среди хвойных всего 4 (Picea abies, P. glauca, Pinus taeda и Pseudotsuga menziesii). Как видите, ДНК транспозонов в обеих группах мало и примерно поровну (около 5%). Ретротранспозонов и у хвойных, и у лиственных значительно больше, чем ДНК транспозонов. При этом у хвойных их доля (около 60%) в два раза выше, чем у лиственных (20-40%).
О чем это говорит. О том, что хвойные не нашли управу на свои ретротранспозоны, не смогли в полной мере показать, кто в геноме хозяин. Большинство МГЭ этого типа в геномах хвойных очень древние. Их накопление происходило методом беспрерывного копирования в течение длительного периода времени, охватывающего десятки миллионов лет. Активность ретротранспозонов в начале жизненного пути хвойных было сугубо положительным явлением. За счет этого явления они обеспечили себе в мезозое быструю эволюцию, большое разнообразие и процветание.
Однако вскоре, к концу мезозоя, процветание сменилось упадком. Не выработав механизма эффективной очистки от «паразитов», геном хвойных оказался сильно отягощен ими, в значительной мере обездвижен и парализован. Дело тут не только в количестве «жира», но и в его качестве. Обычный жир имеет свойство портиться со временем. Геномный тоже. В древних, условно мезозойских ретротранспозонах, к настоящему времени накопилось много вредных мутаций. Поэтому гены, отвечающие за их копирование, часто не работают. Это так называемые геномные пустыни (другой вариант – геномные окаменелости): кладбища когда-то мобильных, но давно уже попорченных и обездвиженных генетических элементов.
У лиственных деревьев ретротранспозонов тоже немало. Но среди них абсолютно преобладают молодые, возрастом не старше нескольких миллионов лет. Цветковые не повторили ошибку, сделанную хвойными. Они выработали более справедливые взаимоотношения с ретротранспозонами: позволили им жить в геноме, оптимизировали скорость их размножения, продолжали их использовать, но при этом научились эффективно удалять лишние, чересчур агрессивные, а также просто выполнившие свою полезную функцию. Это, наряду с другими геномными инновациями, обеспечило цветковым беспрецедентный успех.
МГЭ – самый распространенный тип повторяющейся ДНК. Но не единственный. Копироваться и дублироваться могут также и кодирующие белок последовательности, т.е. гены. Со временем в каждой копии накапливаются свои мутации. Так возникают генные семейства – группы похожих генов с похожими функциями. Эти гены, как правило, располагаются в одной хромосоме недалеко друг от друга. Число генов в семействе варьирует от нескольких копий до нескольких тысяч копий на геном. Когда выяснилось, что у хвойных огромные геномы, сразу возникла гипотеза, что это связано с более крупными генными семействами. В дальнейшем выяснилось, что очень большая часть копий у хвойных не функционируют. Это так называемые псевдогены.
Уникальных (не повторяющихся) белок-кодирующих генов у хвойных примерно 50 000. Много это или мало? Много. Это чуть ли не мировой рекорд. Однако, скорее всего, по мере дальнейшего исследования их число будет сокращаться. Именно так было с человеческим геномом. Сначала предполагалось, что у человека примерно 100 тыс. генов. Теперь наука остановилась всего лишь на 20 тыс. Кстати, когда генов много, в этом нет ничего хорошего. Это признак примитивности генома, под каждую функцию имеющего отдельный ген. Магистральная линия эволюции в этой ее части такая: за счет тонких механизмов регуляции использовать один и тот же ген для синтеза разных белков. Мы коротенько рассмотрели общие свойства хвойного генома. В следующий раз попробуем коснуться некоторых свойств такого вопроса: насколько и чем отличаются геномы разных семейств в подклассе хвойные.