Я приступаю к довольно сложной теме. Метод максимальной парсимонии (экономии) в филогенетике - это тема, которую легко понять поверхностно, но гораздо сложнее осознать глубоко. И, как всегда и бывает, поверхностное понимание чревато ошибочными выводами. Например, парсимонию можно спутать с дистантными методами.
Дистантные методы (метод максимального сходства) - это методы, с помощью которых кодируются состояния для каждого таксона, и потом считается общее количество сходных состояний между таксонами, и на этой основе строится дерево дистанций (или максимального сходства). Это используется как для морфологических, так и для молекулярных данных, но ни там, ни там это не является мэйнстримом.
Это вполне интуитивно понятная методика. Если два организма больше похожи друг на друга, чем на третий, то вполне логично предположить, что первые два более близкородственны. Чем вам не экономия? Но эта логика ошибочна.
Представьте себе акулу, костную рыбу и кошку. Акула и костная рыба больше похожи, чем костная рыба и кошка, однако костная рыба филогенетически ближе к кошке. Как мы это узнали? У костной рыбы и кошки - костный скелет, а у акулы - хрящевой. Костный скелет - это состояние, при котором требуется существенная сложная перестройка работы генов. Если мы предположим, что акула ближе к костной рыбе, то нам надо также принять одно из двух следующих допущений:
1) костный скелет произошел дважды, у костной рыбы и у кошки независимо.
2) костный скелет - это предковое состояние, и по какой-то причине у акулы он стал опять хрящем.
И то, и другое объяснение требует больше допущений, чем допущение, что костный скелет произошел один раз, и его носители стали такими успешными, что в итоге создали множество разнообразных жизненных форм. Именно это и подразумевается, когда мы говорим о максимальной парсимонии.
Возможно (а, может и нет), какой-нибудь специалист по позвоночным мне возразит, и скажет, что костный скелет - это древнее преобразование. Если, мы начнем перебирать вообще все признаки, которые есть у акул, костных рыб и кошек, то может вполне оказаться, что дистантные методы не противоречат выводами парсимонии, и между кошкой и рыбой действительно больше общего сходства.
Ок, тогда представим себе другой пример. Допустим, какая-то популяция какого-то вида летающих насекомых попадает в новую среду обитания. Допустим, ее переносит на маленький остров с сильными ветрами, где полет снижает вероятность выживания, потому что крылатые насекомые будут пытаться летать, и их будет сносить в море, где они будут гибнуть. Таким образом, отсутствие полета будет поддерживаться естественным отбором. Допустим, у каких-то особей произойдет мутация, принципиально важная для полета, и их качество полета станет хуже или эта возможность совсем пропадет. Со временем, эта популяция эволюционирует в какой-то вид, который отличается от того, от которого она изначально произошла. Возможно, у нее редуцируются или вовсе исчезнут крылья, а такие преобразование влекут за собой и другие морфологические и физиологические изменения. Например, очень часто у бескрылых клопов конвергентно схожая квадратная или прямоугольная форма груди, тогда как у крылатых она трапециевидная, и это прослеживается даже в пределах одного вида, когда есть и полнокрылые, и короткокрылые формы.
Те первые нелетающие особи все еще будут иметь крылья, просто перестанут их использовать. Допустим, в какой-то момент на острове живет и нелетающий вид с крыльями и нелетающий вид без крыльев. По общему сходству нелетающий вид с крыльями будет ближе к материковым летающим формам, чем к своим бескрылым собратьям, потому что единственное, что у них есть другого - это мутация, которая усложнила им полет или вообще его запретила. Но именно от них произойдут бескрылые формы, так что эволюционно они будут ближе к бескрылым потомкам, чем к крылатым предкам.
Вот как будет выглядеть реконструкция, сделанная методом максимального сходства:
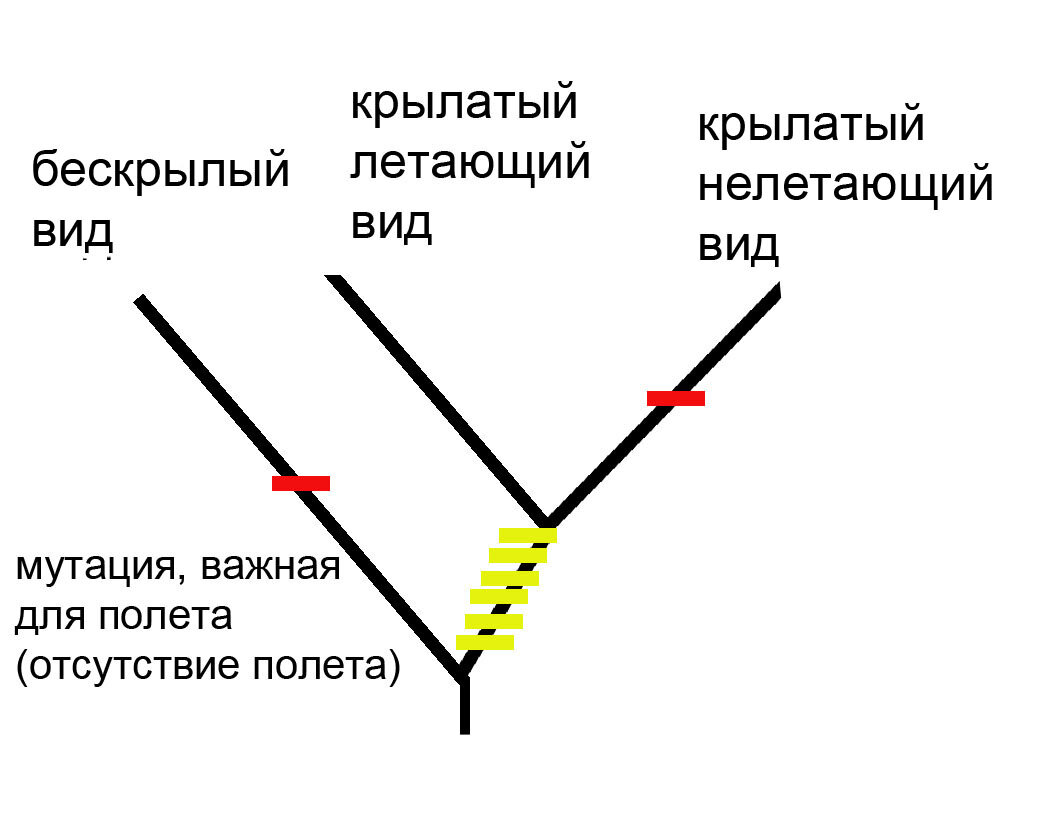
Метод максимального сходства просто сложил все признаки, общие для крылатых видов, и получилось, что они ближе друг к другу. Очевидно, что это неправильная реконструкция, потому что полет редуцировался всего один раз, а на этой реконструкции это произошло дважды (красный признак).
Вот что покажет нам метод максимальной парсимонии:
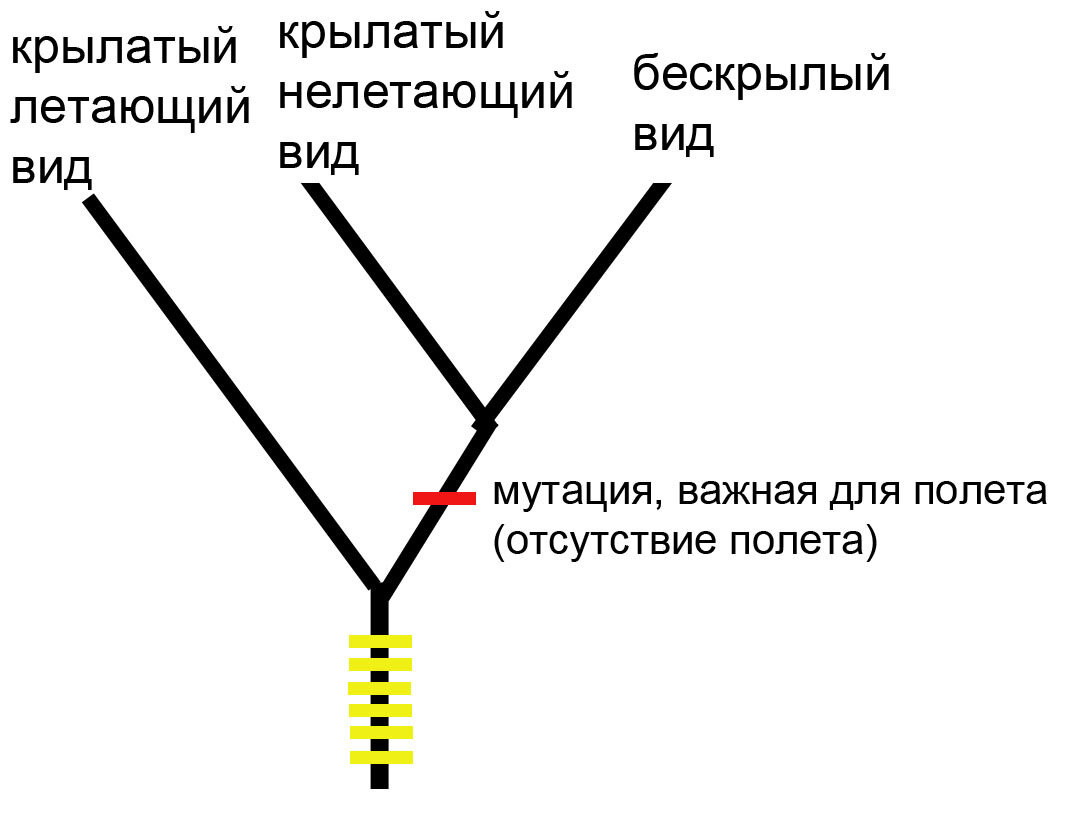
В данном примере это то, что соответствовало бы реальности. Все желтые признаки, которые объединяют крылатый летающий и крылатый нелетающий виды произошли у их общих предков, и на реконструкцию топологии в данном случае не влияют. Конечно, сейчас это просто иллюстрация, нам потребуется много внешних групп и надо будет правильно укоренить филогению, чтобы правильно ее поляризовать.
Конечно, в каких-то случаях максимальная парсимония и максимальное сходство будут давать идентичные результаты. Но есть случаи, когда максимальное сходство будет давать закономерные сбои. Например, в случаях, когда в группе потомков есть формы более похожие на предков, что, как вы понимаете, не редкость, потому что эволюция чаще всего идет постепенно.
И здесь принципиально важны два понятия.
Плезиоморфия - предковый признак. Это признак, который был у предка тех групп, которые мы анализируем, и для анализа несущественен. Например, если мы анализируем филогению насекомых, то у членистоногих организмов, кроме большинства насекомых, крылья отсутствуют, и это не сближает их всех вместе
Апоморфия - продвинутый признак, который сближает какие-то группы. Например, наличие крыльев - это сложный признак, и сложно представить, что он произошел более одного раза. Однако не всегда очевидно, что является апоморфией, а что - нет. К примеру, редукция крыльев - это более простой процесс, и могло происходить в истории насекомых более одного раза. Задача метода максимальной парсимонии выявить апоморфии на основе анализа многих признаков.
Самый очевидный аргумент против парсимониального анализа состоит в том, что в природе бывает всякое, она не обязана действовать экономно. И иногда более сложные допущения оказываются в итоге ближе к реальности.
Во-первых, философия филогенетического анализа состоит в том, что мы работаем с допущениями. Методы, в которых мы применяем эти допущения, - это всего лишь инструменты, с помощью которых мы пытаемся описать и осознать этот мир, и их результаты не выдолблены в камне на века. Это справедливо и для всех остальных методов построения филогений (и вообще для всей науки!). Всякую филогению, неважно, на каких данных она построена, надо рассматривать лишь как филогенетическую гипотезу. Если она будет повторяться в разных исследованиях с разными данными и таксонами, тогда уже можно говорить о том, что, скорее всего, так и было.
Во-вторых, парсимония - конечно, имеет методологические недостатки, но в методах, которые используются для обсчета молекулярных данных, часто используются вероятности. Но чем вероятности лучше, чем парсимония, учитывая, что мы сами задаем начальные параметры анализа?
В-третьих, у нас матрица состоит из большого количества признаков, и учитывается совокупный результат от всех признаков. Это приводит к тому, что некоторые отношения у нас с высокой поддержкой (поддержаны большим количеством признаков), а другие - с низкой (поддержаны малым количеством признаков). И те, что с низкой имеют меньшую достоверность.
Однако, метод максимальной парсимонии, действительно, не рекомендуется использовать для обсчета молекулярных последовательностей Но к этому я вернусь, когда уже буду обсуждать молекулярные данные.
Автор парсимониального анализа - Вилли Хенниг (Willi Hennig), немецкий биолог. В 1950 году у него вышла книга Grundzüge einer Theorie der phylogenetischen Systematik, которая в 1966 году была переведена на английский язык как Phylogenetic Systematics. В те времена эта работа произвела фактически революцию в мышлении биологов, именно потому что он переключил образ мышления от максимального сходства на апоморфии. Я признаюсь, что не читала ни первоисточник, ни перевод. На данный момент, эта книга примерно тоже самое, что "Происхождение видов" Дарвина. Ее историческое значение огромно. Стоит ли ее читать? Я думаю, что ответ на этот вопрос каждый должен принять для себя сам. Конечно, для более глубокого понимания любого вопроса надо прочитать как можно больше литературы и применить это на практике (и осознать результат). Мое мнение - лучше начать с более современной литературы, где методы парсимонии будут освещены применительно к другим филогенетическим методам и современным алгоритмам обсчета.