Допустим у нас уже выбраны внутренняя и внешние группы, и также таксон, на который мы будем укоренять дерево. Что делать дальше? Сначала разберем морфологию. Зачем вообще в наш век быстро развивающейся геномики учить морфологическую филогенетику я объяснила здесь.
Безусловно, чем лучше вы знаете морфологию изучаемой группы, тем лучше.
Но и кодирование признаков тоже может быть важно.
Для построения морфологических филогений чаще всего используют дискретные признаки. Те признаки, которые нельзя разбить на четкие состояния чаще всего откидывают. Однако не всегда, и есть способы в некоторых случаях их тоже использовать, например, размеры какой-то структуры. Но давайте пока поговорим о дискретных признаках.
На примере выдуманных мною существ пятиногов я разберу основные принципы кодирования состояний признаков.
Давайте для начала определимся, что такое «признак», а что такое «состояние признака». Признак — это сущность, то есть идея, а состояние признака — это его конкретное проявление. Состояние признака в филогенетическом понимании более реальное понятие, чем сам признак.
Например, в нашем контексте «цвет шерсти» - это признак, а его состояние — черная, белая, рыжая, то есть конкретное проявление. Признаком также может быть присутствие/отсутствие. Например, если мы захотим закодировать присутствие/отсутствие крыльев, то само по себе это будет признаком, а уже присутствие или отсутствие крыльев у конкретного вида — это состояние признака. То же самое и в молекулярной филогенетике. Определенная позиция в последовательности — это признак, а конкретный нуклеотид в этой позиции или его отсутствие — состояние признака.
Есть два универсальных правила.
1. Чем больше признаков, тем лучше. Логично, что чем больше у нас информации, тем выше вероятность, что у дерева будет высокое разрешение и поддержки.
Тут не все так просто. Иногда бывает так, что признаков много, но это не помогает. Какие тут могут быть проблемы?
- много признаков с уникальными состояниями. Они не несут информации важной для филогении (см. ниже).
- Признаки закодированы некорректно. Например, если крылья птеродактиля и птицы закодировать одним состоянием, это только сделает результаты филогении хуже, потому что эти структуры по-разному устроены.
- Закодированы признаки, которые очень пластичны и легко изменяются в истории группы. Это обычно касается опушения, всяких шероховатостей на теле, размерах.
- Закодированы признаки, которые очень вариабильны внтури вида. Это обычно размеры тела, окраска... Допустим, мы кодируем цвет, и у нас три состояния (0) белый, (1) желтый, (2) коричневый. Во внутренней группе у большинства видов встречается как минимум два состояния. В таком случае признак скорее добавит путаницы в филогению.
2. Минимизировать количество отсутствующих знаков, которые кодируются как ? и - Разницу между ними не скажу, и вроде как на уровне анализа ее нет. Логически понятно, что чем больше у нас отсутствующих данных, тем меньше разрешение и поддержки у результирующей филогении. Однако даже это правило в некоторых случаях будет нарушаться (см. ниже.
Вернемся же к нашим пятиногам. Вот они.
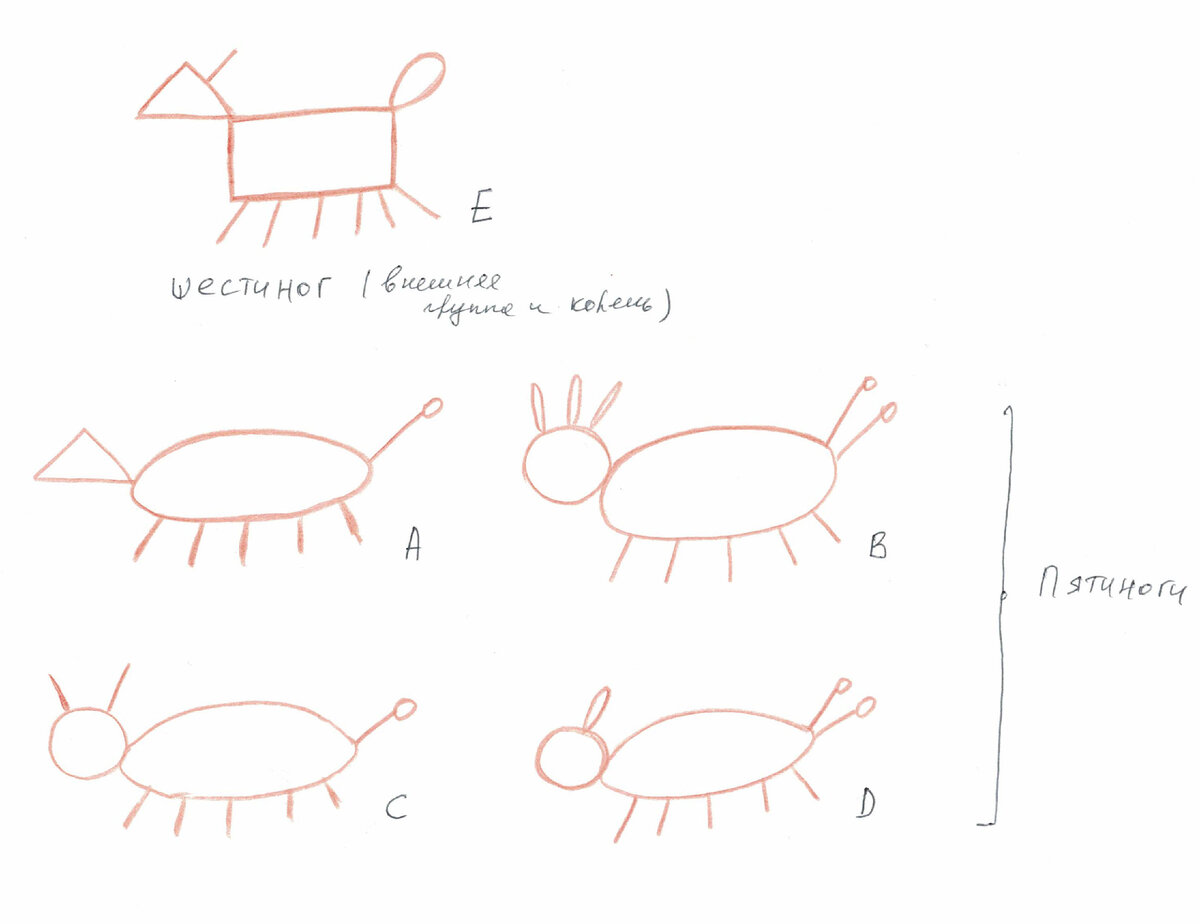
У меня четыре пятинога, и в качестве аутгруппы и корня выступает шестиног.
Нет абсолютных правил для всех случаев, как кодировать признаки. Я привожу матрицу, как бы сделала я.
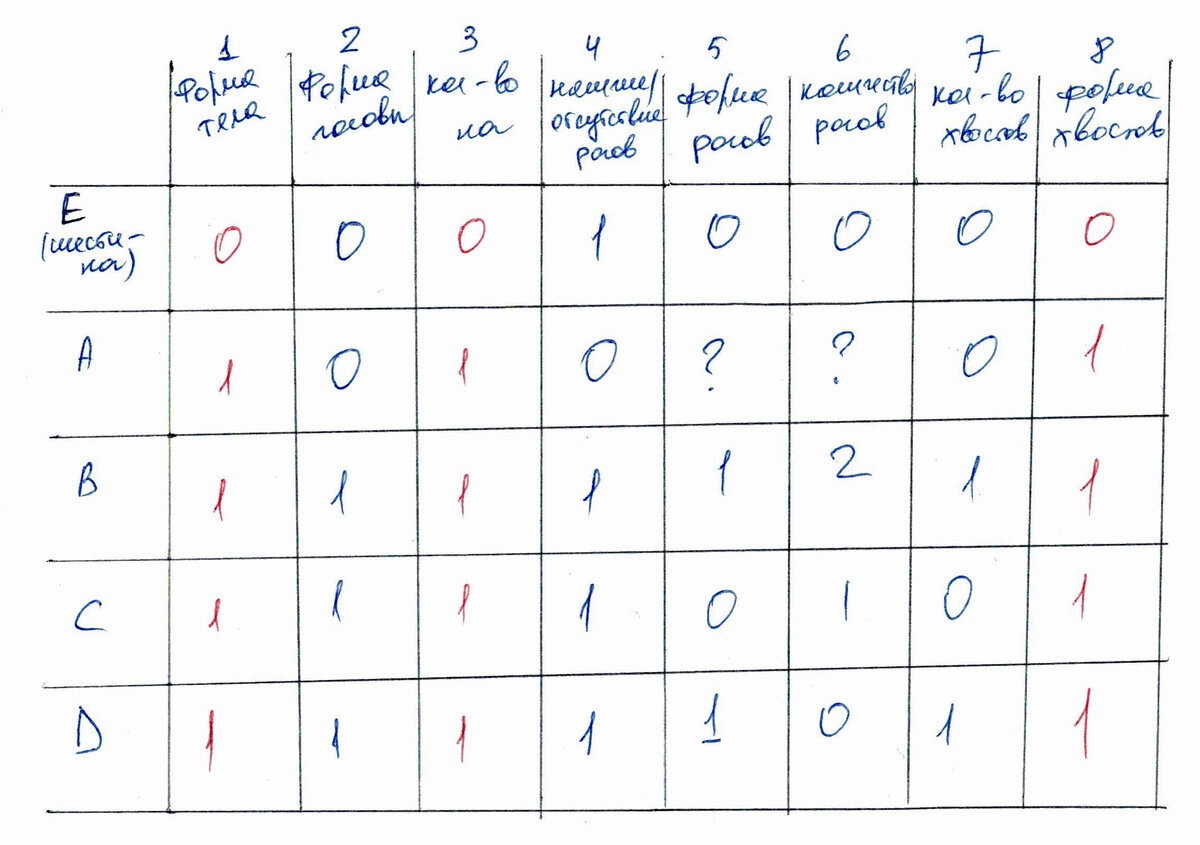
Состояния записываются вот так:
1) Форма тела: (0) прямоугольная, (1) овальная.
2) Форма головы: (0) треугольная, (1) круглая.
3) Количество ног: (0) 6, (1) 5.
4) Наличие/отсутствие рогов: (0) отсутствуют, (1) присутствуют.
5) Форма рогов: (0) палочки, (1) овалы.
6) Количество рогов: (0) 1, (1) 2, (2) 3.
7) Количество хвостов: (0) 1, (1) 2.
8) Форма хвостов: (0) овал, (1) палочка с кружочком на вершине.
Первое, что надо обсудить - это уникальные состояния. Признаки 1, 3, 4, 8 ничего не несут для филогении, потому что они не сближают никакие таксоны . На первый взгляд кажется, что состояние 1 в случае признаков 1, 3, 8 объединяет пятиногов, но это не так. Таксоны группируются друг с другом только относительно других групп таксонов, но относительно одного таксона с каким-то уникальным состоянием,они группироваться не могут. Объединять пятиногов это состояние могло бы, если бы у нас был бы еще один таксон, скажем, с прямоугольной формой тела. Тогда E и этот таксон объдинялись бы этим состоянием, а таксоны A-D относительно них— овальной формой тела.
Чтобы лучше понять эту мысль, представьте себе динозавра, у которого появилось продвинутое состояние - крылья. Делает ли всех остальных динозавров этот факт ближе друг к другу? Очевидно, что нет. Потому что динозавр с крыльями эволюционировал по определению от какого-то динозавра без крыльев. Динозавр без крыльев, произошедший от того же предка, ближе к динозавру с крыльями, чем ко всем остальным. Однако если у нас есть два динозавра с крылями, то уже есть подозрение, что наличие крыльев их может объединять относительно других динозавров без крыльев.
Но если, наоборот, уникальное состояние — это не продвинутое состояние, а, наоборот, предковое? Тогда по логике другое состояние должно сближать те таксоны, у которых оно есть. Но для анализа это ничего не меняет, потому что мы сможем понять, какое состояние предковое, только если поляризуем признаки уже после анализа, как результат укоренения. То есть буквально скажем, что вот этот таксон - корень, и скорее всего его состояния предковые. Во время анализа программа об этом не знает, она строит неукорененную филогению и все состояния расценивает как равноценные. Впрочем, даже если состояние признака есть только у корня, это вовсе не значит, что это состояние предковое. Таксон, на который мы укореняем, также может нести уникальное состояние. И протестировать это мы сможем, только включив больше таксонов с таким же состоянием. Про укоренение написано подробнее здесь.
Это будет еще более очевидно, когда мы подойдем к вопросу о том, как строится филогения.
Таким образом, на самом деле в приведенной матрице признаки 1, 3, 4, 8 для филогении смысла не имеют.
Продолжение по ссылке