Аннотация: В научной статье автор рассматривает ход химико-биологической эволюции, в ходе которого произошло формирование универсального биологического энергоносителя АТФ. В ходе работы, рассматриваются химико-биологические, геологические и физические процессы, идущие в основании процесса формирования АТФ и его производных.
Ключевые слова: АТФ, фосфорилирование, аденозинтрифосфат, универсальный энергоноситель, фосфоангидридные связи, полифосфат
PS: статья опубликована 07.06.2022 года; ссылки на источники находятся в конце статьи.
Энергия – это ключевое понятие, вокруг которого курсирует в принципе все что мы знаем о мире физическом и о живом мире, в частности. Для любого процесса на Земле требуется энергия, для протекания любых биохимических процессов в живых организмах. Существует огромное многообразие различных конечных потребителей энергии в организме и, разумно предположить, что она должна в чем-либо хранится или запасаться.
В случае с живыми организмами долгое по настоящим меркам время, ученые уверенно считали, что единственная возможная разновидность конвертируемой энергетической валюты – это молекула аденозитрифосфата или сокращенно АТФ. Это верно лишь отчасти. Внимательно следите за ходом моей мысли.
Молекула АТФ схематично состоит из 3 частей: аденин, рибоза и три фосфатные группы (рис. 1).
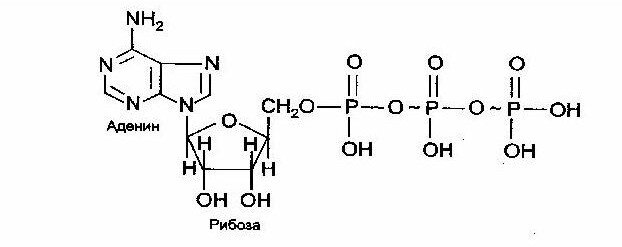
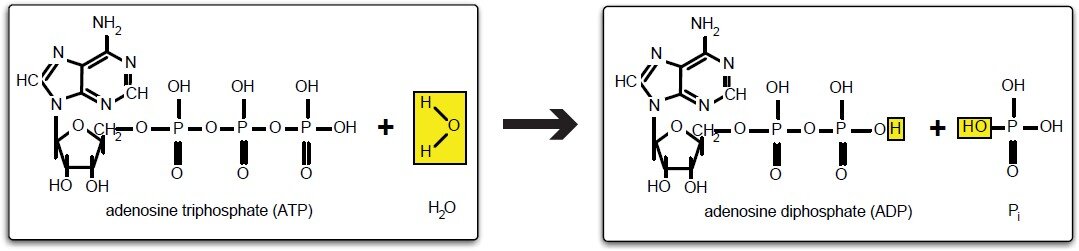
Собственно, высвобождение энергии из этой молекулы происходит при разрыве химических связей, которые связывают фосфатные группы (рис.2). Такие связи называют макроэргическими или иными словами высокоэнергетические, их принято обозначать следующим знаком “~”. Т.е. структуру аденозинтрифосфата можно еще обозначить как:
А - P - Ф ~ Ф ~ Ф
Если обратиться к классической химической терминологии, то такие макроэргические связи между двумя фосфатными группами с отщеплением воды будут называться фосфоангидридными. Сейчас мы рассматриваем самый распространенный тип высокоэнергетической связи, но на самом деле они достаточно многочисленны. Дело в том, что источником энергии в АТФ служат исключительно фосфоангидридные связи и никакие иные. Стоит также упомянуть, что с прагматической точки совершенно не так уж и важно к чему именно эти связи будут прикреплены. Это может быть не аденин, а, например, с равным успехом и гуанин, и урацил. Энергетический эквивалент у них практически не отличается и все они активно участвуют в биохимических процессах: гуанозинтрифосфат (ГТФ) используется при синтезе белка, а уридинтрифосфат (УТФ) – при формировании клеточных стенок у бактерий.
А что произойдет, если мы лишим фосфаты, связанные фосфоангидридными связями их тела в лице нуклеозида? Смогут ли тогда такие полифосфатные цепочки быть энергетически активными в клетке и участвовать в биохимических процессах? Как уже давно было доказано, ответ однозначно положительный. Оказалось, что существуют такие бактерии, у которых полимерные цепи из фосфата активно участвуют в обмене углеводов по точно такому же принципу, как АТФ [1, c. 283-295] . Была досконально изучена одна интереснейшая химическая реакция, которая называется фосфорилирование глюкозы (рис.3).
Фосфатную группу буквально прикрепляют к молекуле глюкозы, и в результате мы получаем глюкозо-6-фосфат. Это важнейшая биохимическая реакция, с которой начинается усваивание глюкозы в энергетическом обмене. У подавляющего большинства организмов, как это не удивительно, источник фосфата представлен исключительно в виде молекулы аденозитрифосфата (АТФ). Но есть важное исключение, - некоторые бактерии используют для этой реакции не АТФ, а обыкновенный полифосфат.
Как отмечает в своей работе Игорь Кулаев (советский и российский биохимик), проводя совместные исследования с лабораторией М. Шимоны: “было установлено, что реакция использования высокомолекулярных полифосфатов вместо АТФ для фосфорилирования глюкозы присутствует и активно функционирует только у небольшого количества эволюционно древних бактерий (пропионовые бактерии, микрококки, тетракокки, и некоторы е другие), принадлежащих только к одной небольшой веточке эволюционного древа. А у более современных групп бактерий и других организмов вместо высокополимерных полифосфатов используется только АТФ” [2]
Из этих достоверно подтвержденных неоднократно наблюдений могут следовать соответствующие выводы о том, что биохимические реакции с использованием чистого полифосфата это не что иное, как эволюционно древний способ использования энергоносителей. А те немногие виды бактерий, которые до сих пор используют полифосфатные цепи, а не эволюционно новый АТФ – являются яркими представителями ископаемых живых систем. Наглядно продемонстрируем, чем отличаются реакции фосфорилирования глюкозы предположительно древних организмов с использованием полимеров фосфата и эволюционно новых реакций с использованием АТФ (рис.4, рис.5):
В настоящее время любые процессы обмена с глюкозой всегда начинаются исключительно с реакции активирования глюкозы посредством использования аденозитрифосфата (рис. 2)
Есть несколько весьма весомых доводов в пользу теории о том, что именно полифосфаты с макроэргическими связями были непосредственными эволюционными предшественниками АТФ.
- Первый важнейший довод говорит о том, что времена, когда активно происходило зарождение жизни, сопровождались невероятными по масштабу тектоническими и вулканическими процессами. Это наглядно показывают исследования, проведенные Ю. Ямагата, Х. Ватанабе и их коллегами, опубликованные в статье “О значении вулканической активности для пребиотической эволюции” [3, c. 516-519] В следствии чего происходила конденсация летучих соединений фосфора, которые в большом количестве содержаться как раз в вулканических газах.
- Во-вторых, при анализе древних горных пород, в частности, архейских коматиитов и метаморфизованных базальтов в Гренландии, были смоделированы процессы восстановления неорганического фосфора в доступный для биологических форм. Было показано, что на т.н. ранней Земле также была сильно повышенная грозовая и метеоритная активности (рис.6). Считается, что эти факторы совместно участвовали в образовании и накоплении пребиотического фосфора в период с 4,5 до 3,5 млрд лет назад [4].
Как мы можем видеть, очень вероятно, что чистые фосфатные полимеры вероятно могли быть древними формами энергоносителей для ископаемых форм живых организмов.
Переходя далее, вспомним, что фосфатные группы в современных организмах неизменно прицеплены к масштабной молекулярной структуре, к т.н. носителю. В АТФ эту функцию выполняет нуклеозид аденина с рибозой (рис. 1). Как мы уже замечали ранее, в виде тела для фосфатных групп вполне может подойти и уридин, и гуанозин, и даже цитидин. При этом в энергетическом эквиваленте не произойдет никаких критически важных изменений. Но почему в таком случае, универсальной молекулой энергоносителем стала именно аденозинтрифосфорная кислота, а не какое-либо иное похожее соединение, ведь, как мы видим, выбор действительно есть. Для этого по всей видимости были причины.
Если мы внимательно рассмотрим графические формулы пуриновых и пиримидиновых оснований (рис.7), то увидим малую, на первый взгляд особенность, у аденина нет атомов кислорода и именно это свойство его ярко выделяет на фоне прочих азотистых оснований, которые распространены наиболее широко.
Во времена, когда формировалась жизнь в ее фундаментальном начале, свободного кислорода в атмосфере было критически мало или не было вообще (рис. 8). Что позволяло синтезироваться аденину очень легко, в отличие от других азотистых оснований, в которые входят атомы кислорода.
Также, стоит вспомнить про одну интересную взаимосвязь, что в эпоху, когда жизнь только-только начинала формироваться, на Земле еще не сформировался озоновый слой в силу того, что озон состоит из трех атомов кислорода, а как мы уже выяснили, свободного кислорода в атмосфере еще не практически не было. То есть солнечная радиация попадала прямо на земную поверхность совершенно беспрепятственно, а это в свою очередь могло приводить к интересным реакциям с участием аденина, например, к реакции фотофосфорилированию. Если кратко, то это процесс синтеза АТФ из АДФ (аденозиндифосфат) при участии солнечного излучения. Нуклеотиды же с другими азотистыми основаниями таким свойством не обладают, это было доказано во множестве экспериментальных наблюдений и аналитических моделях. Такое исключительно удивительное свойство аденина могло запросто стать причиной его переизбытка в древних эпохах, что и ознаменовало его успех в эволюционной гонке за право быть универсальным энергоносителем для биологических организмов [5].
Также АТФ является далеко не самым высокоэнергоемким соединением из всех тех, что доступны в живой природе. Как альтернативные варианты можно с успехом рассмотреть глицерофосфат или же не менее интересное соединение фосфоенолпируват. Оба эти соединения содержат в себе фосфатные группы, но при этом по структуре не схожи в аденозитрифосфатом. Но аналогично с АТФ они могут путем обыкновенного гидролиза выделять энергию, отдавая молекулу фосфата. Вычислив количественные значения выделяемой энергии, мы получим подтверждение об отсутствии у АТФ превосходства по энергоёмкости:
На основании анализа вышеизложенных данных, мы вправе сделать некоторые выводы: по всей видимости, длинные полифосфатные цепочки и вправду могли быть эволюционными предшественниками для современных энергоносителей в виде АТФ. Особо серьезных препятствий к этому не было. АТФ же в свою очередь, будучи не самым энергоемким и уникальным соединением, выдержало давление биологическим отбором и стало универсальной энергетической составляющей. Также стало ясно и то, что коэффициент энергетической емкости, вероятно, не был движущим фактором, по которому следовала биологическая эволюция при формировании живых организмов, гораздо важнее было то, чтобы то или иное соединение могло успешно связываться с другими молекулами и успешно участвовать в биохимических процессах, другими словами, чтобы соединение было энергоэффективным, так оно и вышло.
Были приведены множественные примеры закономерных процессов, которые потенциально могли сопутствовать постепенному становлению АТФ в современном статусе универсального энергоносителя. Большая часть из них имеет твердую теоретическую и практическую базу, достоверность которых проверена в экспериментальных условиях, а также в ходе математического моделирования.
Библиографический список:
1. Shymona M., Ostrowski W. Inorganic polyphosphate glucokinase of Mycobacterium phlei // Biohimica et Biophysica Acta (BBA), Specialized Section on Enzymological Subjects, 1964, V. 85, №2, 283-295.
2. И.С. Кулаев “Неорганические полифосфаты”// Библиотека статей URL: http://www.gendna.ru/arhiv/biology/polyphosphate.html
3. Yamagata, Y., Watanabe, H., Saitoh, M. et al. Volcanic production of polyphosphates and its relevance to prebiotic evolution. Nature 352, 516–519 (1991).
4. Hess, B.L., Piazolo, S. & Harvey, J. Lightning strikes as a major facilitator of prebiotic phosphorus reduction on early Earth. Nat Commun 12, 1535 (2021)
5. Скулачев В.П. Эволюция биологических механизмов запасания энергии // Соросовский образовательный журнал. 1997. №5.
6. Солвей Дж. Г. Наглядная медицинская биохимия. Учебное пособие. — М.: ГЭОТАР-Медиа, 2020. — 168 c.
7. Уилсон К., Уолкер Дж. Принципы и методы биохимии и молекулярной биологии. — М.: Лаборатория знаний, 2021. — 848 c.