Уважаемый читатель, сначала вам предстоит ознакомиться с критикой:
https://yadi.sk/i/VBb9PKbZvWf-vQ
Как видите, написано очень много, в основном переливая из пустого в порожнее. Попробуем вам помочь и выделить суть критики в нескольких предложениях.
Критик считает, что модель поэтапного рекрутирования из-за накопления метаболитов неверна на основе эксперимента: https://www.instagram.com/p/BnnjZtNHo67/
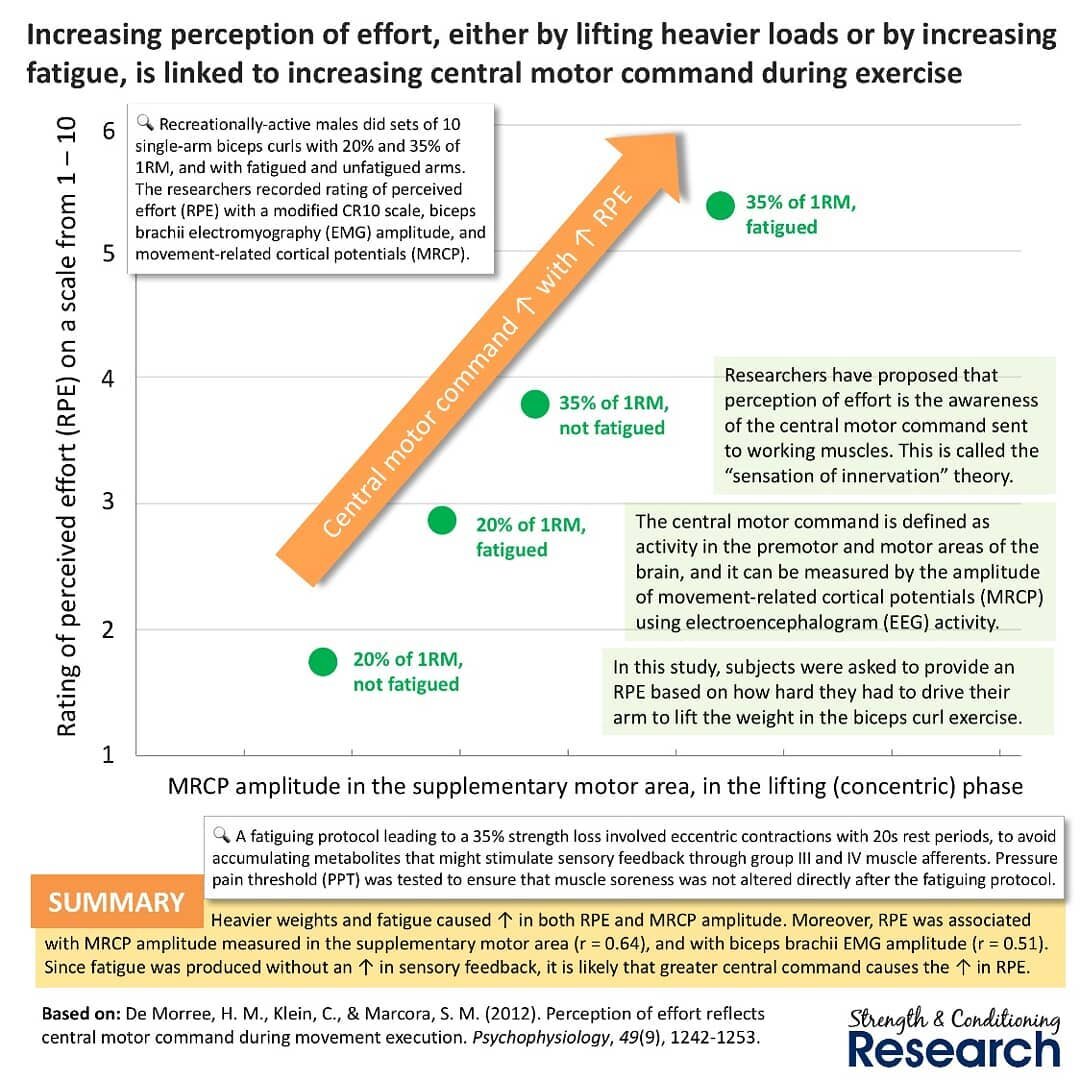
в котором подопытные выполняли сгибания руки в локте с отягощением 20% и 35% (с темпом: 1 секунда подъем и 1 секунда опускание) на «свежую» и с предварительным утомлением. Предварительное утомление приводило к 35% потере силы и заключалось в эксцентрических сокращениях с 20 секундным периодом отдыха, чтобы исключить образование метаболитов. И в том и другом случае было повышение сигнала из ЦНС, что говорит о том, что роль метаболитов в поэтапном рекрутировании не имеет значения или является только частным случаем. А значит считавшаяся ранее традиционная модель гипертрофии не соответствует действительности.
В традиционной модели считается, что мышцы гипертрофируются от упражнений с использованием больших отягощений в результате механического напряжения, а в случае с малым сопротивлением – в результате накопления метаболического стресса. В то время как по мнению критика (а его выводы опираются на теорию Криса Бердсли), при использовании малых отягощений происходит поэтапное рекрутирование и замедление скорости сокращения в конце подхода до отказа, что приводит к активации высокогопороговых двигательных единиц и повышению механического напряжения в этих волокнах, что и приводит к гипертрофии.
Очень странным выглядят такие выводы от человека, который, по его словам, досконально изучил спортивную адапталогию,

так как данный эксперимент только подтверждает её. А то что традиционная модель ошибочна В. Н. Селуянов понял еще в начале 80-х годов прошлого столетия…
А теперь по порядку.
Модель поэтапного рекрутирования на основе накопления метаболитов действительно ошибочна и это достаточно неплохо показано в приведенном ранее эксперименте. Модель поэтапного рекрутирования в спортивной адапталогии выглядит следующим образом. Рекрутирование происходит в соответствии с правилом Ханнемана. Рекрутирование новых мышечных волокон осуществляется не только в результате утомления МВ, т.е. накопления ионов водорода и снижения силы сокращения, но и ослабления МВ, т.е. перехода на другой вид энергообеспечения, сопровождающийся падением мощности.
Для более полного понимания следует рассмотреть это на условном примере поэтапного рекрутирования при небольшой нагрузке.
При использовании некоего малого сопротивления сначала в работу будут включены только ОМВ, но после 15-20 секунд работы на запасенном АТФ и креатинфосфате, мощность снизится, примерно, до 30-50% от начального уровня, в результате перехода на аэробный гликолиз. Для поддержания мощности, ЦНС повысит частоту сигнала и рекрутирует часть ПМВ, мощность которых также снизится через 15-20 секунд работы, далее в ход пойдут ГМВ до исчерпания двигательных единиц. Но если ОМВ и ПМВ после перехода на аэробный гликолиз продолжают работу и выдавать 30-50% своей мощности, то в ГМВ после 15-20 секунд работы начинается лавинообразное нарастание ионов водорода, которые с каждой секундой всё больше препятствуют сокращению и через 60 секунд ГМВ не сможет сокращаться вообще.
Это лишь очень примерное описание того, что происходит в мышце, так как МВ начинают поэтапно рекрутироваться, постепенно повышая напряжение, т.е. сначала мышца начинает работу в зубчатом тетанусе и затем переходит на сокращение в гладком тетанусе. Это позволяет сглаживать силу, в противном случае, она росла бы рывками.
О других вариантах происходящих при поэтапном рекрутировании будет изложено в последующих статьях.
Так что данный эксперимент только подтверждает данные спортивной адапталогии.
Что касается эксцентрического режима сокращения, то траты энергии в нём зависят от времени выполнения. Трат АТФ на разрыв актин-миозиновых мостиков не будет, если расслабить мышцу и позволить конечности упасть под действием силы тяжести, в данном случае актин-миозиновые мостики вообще не будут образованы, а сопротивление будет совсем минимальным только за счет упругих компонентов. В случае же очень быстрого удлинения активной мышцы актин-миозиновые мостики будут разорваны механически практически без трат энергии. Если же эксцентрика не будет очень быстрой, то молекулы АТФ успеют отщеплять намного больше мостиков, остальная небольшая часть будет разорвана механически.
Поэтому уже рассмотренное в предыдущих частях исследование на основе прыжков в глубину (https://www.ncbi.nlm.nih.gov/pubmed/15954988) – не может служить доказательством отсутствия трат энергии. Классическое выполнение прыжков в глубину на Западе выполняется следующим образом: атлет, сходя с возвышенности должен приземлиться на опору и «поглотить энергию», образованную падением тела, за счет упругих структур: сухожилий и мышц. При данном варианте исполнения спортсмен проводит в полуседе около 1-2 секунд (зависит от высоты тумбы), тем самым тратя энергию, как и при вставании. А так как объем спрыгиваний составляет 100 повторений подряд, то в данном случае будут не только траты АТФ, но и небольшое образование ионов водорода. Поэтому данный эксперимент не может служить подтверждением высказываний оппонента, тем более, что данных об утомлении в данном эксперименте нет (по крайней мере в аннотации).
Хорошим подтверждением сказанного выше и является ранее приведенное оппонентом исследование: https://pbs.twimg.com/media/DrdkkZ3W4AA9KAa.jpg
Из которого видно, что рост лактата был в случае «красного» протокола (50% от ПМ, 5 секунд в уступающем и 1 секунда в преодолевающем режиме). Если сравнить с данными полученными в «синей» группе, то мы можем наблюдать очень небольшое повышение лактата: чуть более 5 ммоль/л (на пике) в «красной» группе при 10 секундах работы в концентрическом режиме и 50 секунд в эксцентрике, и чуть менее 6 ммоль/л (на пике) в «синей группе» при 50 секундах концентрического режима сокращения и 10 секунд эксцентрического. Что и говорит о правильности наших выводов.
Теперь коснемся еще нескольких исследований и утверждений.
Очень странно, что критики спортивной адапталогии, некоторые положения которой базируются на математическом моделировании проходящих процессов, оппонируют, используя исследования, которые также базируются на математическом моделировании, только используют при этом достаточно простую модель, основанную на статистических данных (читай: средней температуре по больнице): https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5473583/ Но при всём при этом оно никоим образом не противоречит данным спортивной адапталогии.
Оппонент утверждает, что в конце подхода до отказа рекрутируются высокопороговые МВ и за счет снижения сокращения, в них образуется максимальное количество актин-миозиновых мостиков, что приводит к увеличению механического напряжения, а это в свою очередь приводит к гипертрофии. Но как же быть с ранее приведенным вами экспериментом, в котором наглядно показано, что ММВ тоже гипертрофируются? https://www.ncbi.nlm.nih.gov/pubmed/27174923
Также оппонент утверждает, что только при снижении скорости сокращения – сила сокращения максимальна за счет максимального образования актин-миозиновых мостиков. Да это так, но это в первую очередь касается ММВ, а не БМВ, так как только при медленном характере движения ММВ успеет показать свою максимальную силу, а БМВ смогут сделать это и при более быстром сокращении, так как частота гребков у БМВ выше примерно в 2 раза.
Уважаемые оппоненты и критики, не спешите с написанием подобной критики, не тратьте своё и чужое время попусту. Пишите кратко и по делу.
У всех есть ошибки, это нормально, но их необходимо находить и исправлять. Мы говорим лишь о моделях, а любая модель содержит ошибки (абсолютно любая, просто ошибки бывают разного рода). Вместо нахождения ошибок, вы пока занимаетесь какой-то бестолковой болтологией.
Продолжение возможно...
Вступайте в группу Александра Грачёва: https://vk.com/grachevsport
Будем признательны в помощи по распространению данной статьи!