Почему о цианобактериях так часто говорят?
- Цианобактерии - настоящие космополиты. Разные виды цианобактерий можно встретить и в пресных, и в соленых водоемах, в верхних слоях почвы, на камнях и даже в горячих источниках.
- Цианобактерии - первые обладатели оксигенного, т.е. идущего с выделением кислорода, фотосинтеза.
- Цианобактерии вызвали кислородную катастрофу около 2,4 млн. лет назад и создали современную кислородную атмосферу Земли.
- Согласно теории симбиогенеза, хлоропласты эукариот произошли от предковых цианобактерий в результате первичного эндосимбиоза.
- Цианобактерии способны фиксировать атмосферный азот и переводить его в биологически доступные формы.
- Цианобактерии образуют множество симбиозов с другими организмами: грибами, растениями, животными.
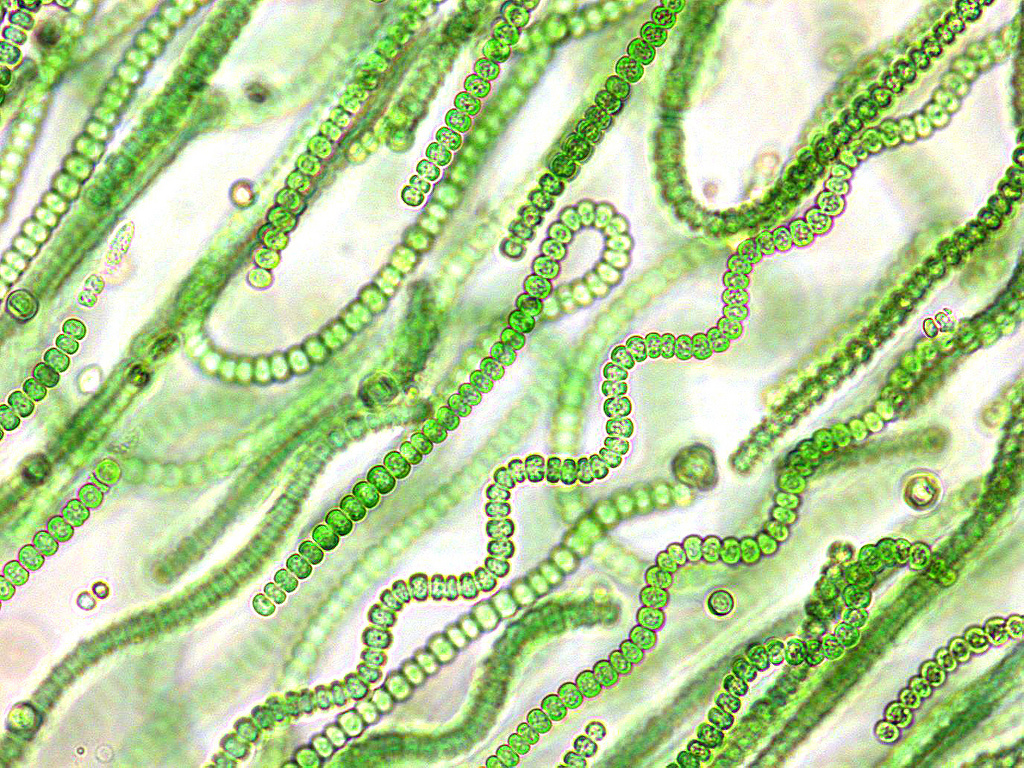
Особенности строения
Цианобактерии являются грамотрицательными бактериями.
Внутренний слой их клеточной стенки состоит из муреина (пептидогликана), при этом у цианобактерий он обычно толще, чем у типичных грамотрицательных бактерий.
Наружная мембрана содержит липополисахариды.
Слизистая капсула включает пектиновые вещества и мукополисахариды. Она выполняет защитную функцию, предотвращает высыхание, участвует в движении (скользящий тип движения).
Генетический аппарат представлен кольцевой ДНК, как у всех прокариот и не окружен мембраной. Это показывает, что цианобактерии - именно бактерии, т.к. долгое время их считали одноклеточными водорослями. До сих пор можно встретить устаревшее название - сине-зеленые водоросли.
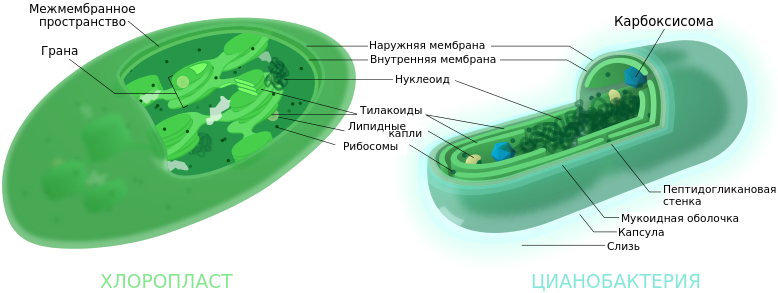
Цианобактерии - фотосинтезирующие организмы, но в отличие от эукариот, у них нет хлоропластов. Вместо этого существует система впячиваний плазмалеммы (плазматической мембраны). Тилакоиды представляют собой уплощенные мембранные мешочки, свободно лежащие в цитоплазме. Они не собраны в граны, как у растений.
На поверхности мембраны расположены фикобилисомы - надмолекулярные белковые комплексы, служащие светособирающими «антеннами» для фотосинтеза.
Фикобилисома имеет форму полусферы, прикреплённой к мембране тилакоида и содержит водорастворимые пигменты-антенны:
- Фикоцианин (синий пигмент)
- Фикоэритрин (красный пигмент)
- Аллофикоцианин.
Пигменты обеспечивают перенос энергии по каскаду фикоэритрин – фикоцианин - аллофикоцианин - хлорофилл a (фотосистема II).
Хлорофилл a является обязательным компонентом фотосистемы. Хлорофиллы b и c у цианобактерий отсутствуют.
Каротиноиды защищают хлорофилл от фотоокисления, особенно у поверхностных видов.
Хлорофилл и фикоцианин обуславливают сине-зеленый цвет бактерий, однако окраска может варьироваться.
Для эффективной фиксации углерода в процессе фотосинтеза цианобактерии имеют карбоксисомы - многогранные однослойные белковые тела, содержащие фиксирующие углерод ферменты, в т. ч. фермент РуБисКО.
CO₂ накапливается внутри карбоксисом, за счет чего РуБисКО работает эффективнее, не тратя энергию на оксигеназную реакцию - фотодыхание.
Предполагается, что карбоксисомы появились в процессе эволюции цианобактерий как адаптация к пониженной концентрации углекислоты.
В воде CO2 диффундирует медленнее, чем в воздухе, а его растворимость сильно зависит от pH. Кроме того, фермент РуБисКО чувствителен к высоким концентрациям кислорода, который является конкурентом за его активный центр.
Обитающие в воде цианобактерии для обеспечения плавучести имеют газовые вакуоли (аэросомы), состоящие из множества белковых цилиндров, проницаемых для газов, но непроницаемых для воды.
Запасные вещества цианобактерий достаточно специфичны:
- вместо крахмала они накапливают цианофициновый крахмал, который в отличие от обычного крахмала, йодом окрашивается не в фиолетовый, а в бурый цвет и по структуре ближе к гликогену животных.
- цианофицин (не путать с цианофициновым крахмалом!) - полипептид из аспарагина и аргинина – обеспечивает запас азота в клетке.
- волютин - внутриклеточные гранулы, состоящие из неорганического полифосфата, составляющие запас фосфора для синтеза АТФ.
- липидные капли.
Многие нитчатые цианобактерии демонстрируют клеточную дифференцировку, что нетипично для прокариот.
У нитчатых форм в трихоме (нити) содержатся следующие виды клеток:
- Вегетативные клетки - обычные клетки, способные фотосинтезировать и делиться.
- Гетероцисты - клетки, способные фиксировать молекулярный азота (N₂ - NH₃). Эти клетки имеют толстую многослойную оболочку - ламинарный слой гликолипидов, препятствующий диффузии газов, лишены фотосистемы II, поэтому не выделяют O₂ и не фиксируют CO₂, при этом их фотосистема I активна, поэтому в ходе циклического фотофосфорилирования они способны получать АТФ. Гетероцисты соединены с соседними клетками микроплазмодесмами, через которые получают малат и глюкозу, отдавая аминокислоты (глутамин).
- Акинеты - очень крупные, заполнены цианофицином и гликогеном клетки, имеющие плотную оболочку необходимы для переживания неблагоприятных условий (холод, высыхание, недостаток фосфора).
- Гормогонии - короткие, подвижные фрагменты трихома, лишенные гетероцист и чехла. Служат для размножения и расселения.
Жизненный цикл
Цианобактери размножаются исключительно бесполым путем - бинарным делением.
Вегетативная стадия представляет собой нить с вегетативными клетками и гетероцистами.
Под действием стресса или снижения уровня азота нить распадается в местах соединения с гетероцистами. Образуются гормогонии.
Гормогоний перемещается по субстрату за счет секреции слизи. Когда гормогоний теряет подвижность и покрывается слизистым чехлом, часть его клеток превращается в новые гетероцисты, и формируется новая колония.
При ухудшении условий вегетативные клетки превращаются в акинеты, прорастая в новый трихом при наступлении благоприятных условий.
Роль в экосистемах
Круговорот углерода
Цианобактерии являются оксигенными фотоавтотрофами. Они фиксируют CO₂ в цикле Кальвина. Карбоксисомы позволяют эффективно работать РуБисКО даже при низких концентрациях CO₂ в воде, обеспечивая «биологический углеродный насос».
Потребляя CO₂ в воде, они сдвигают карбонатное равновесие (HCO₃⁻ - CO₃²⁻), что вызывает осаждение известняка (CaCO₃). Минерал "цементирует" верхний слой бактерий и те отмирают, но сверху вырастает новая колония.
Со сременем формируются минеральные образования с характерными концентрическими карбонатными слоями - строматолиты.
В настоящее время самые известные строматолиты можно встретить в заливе Шарк (Австралия) и в районе Багамских островов.
Круговорот азота
Многие цианобактерии обладают ферментным комплексом нитрогеназой, способным восстанавливать атмосферный азот (N₂) до аммония (NH₄⁺).
Проблема в том, что этот процесс происходит в строго анаэробных условиях, тпоскольку нитрогеназа необратимо ингибируется кислородом.
Для защиты от кислорода разные виды цианобактерий выработали следующие приспособления:
- Пространственное разделение: фотосинтез (выделение O₂) идет в вегетативных клетках, а фиксация N₂ - в гетероцистах.
- Временное разделение: у одноклеточных (не нитчатых) цианобактерий (напр. Cyanothece) нет гетероцист. Фотосинтез идет днем, а азотфиксация - ночью.
- Коллективное разделение: в трихоме без гетероцист (напр., Trichodesmium) часть клеток «отключает» фотосистему II и временно выполняют функции азотфиксации.
При разложении отмерших цианобактерий, азот из органических соединений превращается в доступный для растений аммоний.
Цианобактерии формируют основу морских пищевых цепей (пикопланктон).
Пикопланктон - мельчайшие планктонные организмы, размеры которых варьируются от 0,2 до 2 микрометров (мкм).
Кроме того, цианобактерии - участники множества симбиозов с другими организмами.
В составе лишайников, цианобактерии выполняют функцию фотобионта (фотосинтезирующего организма).
У папоротника Azolla в полостях листьев живет цианобактерия Anabaena azollae, являющаяся поставщиком биодоступного азота. Симбиоз используется на рисовых полях как «зеленое удобрение».
Среди голосеменных саговники образуют симбиоз с цианобактериями в коралловидных корнях.
У покрытосеменных цианобактерия проникает внутрь растения Гуннера через специализированные железистые органы, расположенные в основании листьев.
Оказавшись внутри, цианобактерия проникает в саму клетку гуннеры. Бактерия превращается в гетероцисту, теряя способность к фотосинтезу, но взамен обеспечивая растение азотом.
С губками в бедных азотом тропических водах цианобактерии образуют симбиоз, снабжая азотом.
Однако вместе с тем, цианобактерии могут образовывать на губках биопленки, вызывая их гибель.
Массовая гибель эндемичных байкальских губок вследствие активного размножения нитчатых цианобактерий - одной из главных экологических проблем озера Байкал.
Цветение цианобактерий
Эмассовое размножение бактерий, вызванное прогревом воды выше +20°C, слабой циркуляцией и избытком фосфора. Цвет воды меняется на ярко-зеленый.
Активный рост цианобактерий перекрывает доступ солнечного света, поглощает растворенный кислород и выделяет токсины, что приводит к массовой гибели рыбы и других водных организмов.
Итог
- Устаревшее название цианобактерий - сине-зеленые водоросли.
- Цианобактерии - прокариотические организмы.
- Цианобактерии - космополиты, обитают в различных условиях, включая горячие источники.
- Цианобактерии - первые обладатели оксигенного фотосинтеза.
- Цианобактерии вызвали кислородную и создали современную кислородную атмосферу Земли.
- Хлоропласты эукариот произошли от предковых цианобактерий.
- Цианобактерии могут быть одноклеточными и нитчатыми.
- У нитчатых форм клетки выполняют разные функции - есть фотосинтезирующие и азотофиксирующие клетки (гетероцисты), а также формы для перенесения неблагоприятных условий (акинеты).
- Т.к. цианобактерии - прокариоты, у них нет пластид, но есть светоулавливающие комплексы на мембране - фикобилисомы.
- Для повышения эффективности фиксации углекислоты, у цианобактерий есть карбоксисомы.
- Цианобактерии накапливают углеводы не в форме крахмала, а в форме цианофицинового крахмала, близкого к гликогену.
- Цианобактерии способны фиксировать атмосферный азот и переводить его в биологически доступные формы.
- Цианобактерии образуют множество симбиозов с другими организмами: грибами, растениями, животными.
- Цианобактерии сформировали строматолиты.
- Цианобактерии вызывают цветение воды в жаркое время.