
Наконец я добралась до перевода статьи, на которую так часто ссылалась как на материал с реконструкцией внешнего вида собаки Жохова, так и на сравнение генетического родства различных ездовых пород и популяций собак: DOI 10.1093/gbe/evae190.
Авторы: Tracy A Smith 1), Krishnamoorthy Srikanth 2), Heather Jay Huson 2);
1) Department of Biological Sciences, University of Maryland Baltimore County, Baltimore, MD 21250, USA
2) Department of Animal Sciences, Cornell University College of Agriculture and Life Sciences, Ithaca, NY 14853, USA
Аннотация
Последние научные данные демонстрируют геномную и морфологическую преемственность внутри арктической родословной собак. Здесь авторы используют породу сибирский хаски для изучения геномного наследия северо-восточной евразийской арктической родословной и моделируют глубокую популяционную историю, используя однонуклеотидные полиморфизмы по всему геному. Используя молекулярные часы, откалиброванные с помощью геномов древних собак, они обнаружили, что по крайней мере две различные линии арктических собак существовали в древней Евразии в конце плейстоцена. Это отодвигает в прошлое происхождение ездовых собак на северо-востоке Сибирской Арктики, поскольку люди, вероятно, намеренно отбирали собак для выполнения различных функций и поддерживали размножающиеся популяции, которые пересекались во времени и пространстве, оставаясь в относительной репродуктивной изоляции. У современных сибирских хаски мы обнаружили сложную популяционную структуру, основанную на том, как именно они используются людьми, а также недавнюю интрогрессию европейских пород примерно у половины исследованных собак, участвующих в гонках, умеренный уровень инбридинга и меньшее количество потенциально вредных вариантов в популяциях, подвергающихся строгому селективному отбору по форме и функциям (шоу, ездовые-шоу и гоночные популяции сибирских хаски). Поскольку борьба за сохранение уникальных эволюционных линий при сохранении генетического здоровья породистых собак усиливается, понимание геномной истории для определения политики и наилучших методов управления породой имеет решающее значение для сохранения этих древних линий и их уникальной эволюционной идентичности.
Значение тематики
Ранняя эволюция и генетическое здоровье собак вызвали значительный интерес как у общественности, так и у более широкого научного сообщества. В этом исследовании авторы рассматривают породу сибирский хаски как образец северо-восточной евразийской арктической родословной собак и проводят одно из крупнейших геномных исследований среди всех современных пород. Они реконструировали эволюционную историю этой линии и обнаружили, что современные американские и евразийские арктические ездовые собаки имеют более отдаленное родство, чем считалось
ранее, таким образом отодвинув происхождение ездовых собак на конец плейстоцена. Недавнее проникновение в Западную Евразию, небольшая численность популяции и инбридинг создают проблемы для сохранения этих древних линий.
Введение
Последние достижения в области филогеномики, популяционной генетики, анализа древней ДНК и сравнительной геномики позволяют реконструировать прошлое и глубже проникнуть в происхождение и эволюцию собак. Общепризнано, что собака эволюционировала вместе с популяциями охотников–собирателей в Евразии где-то в позднем плейстоцене от ныне вымершего предка-волка. Хотя до сих пор ведутся споры о том, когда именно, где и сколько раз происходило одомашнивание (Savolainen и др. 2002; Pang и др. 2009; Freedman и др. 2014; Skoglund и др. 2015; Perri и др. 2021), недавние находки показывают, что современные собаки более тесно связаны с древними волками из Восточной Сибири и Китая, чем с волками из Западной Евразии, что указывает на вероятное одомашнивание в Восточной Евразии (Bergström и др., 2022). Однако до половины предков древних и современных западных собак связаны с юго-западными евразийскими волками, что свидетельствует о генетической преемственности от неолита до современных европейских собак (Bergström и др. 2022). Это может свидетельствовать о вторичном одомашнивании или о большом количестве древней примеси западного волка в этих регионах (Bergström и др., 2022). К 11000 лет назад, по крайней мере, три основные родословные собак были хорошо известны: восточноазиатская, западноевразийская (включая европейских, индийских и африканских собак) и арктическо-американская (Frantz и др. 2016; Ní Leathlobhair и др. 2018; Bergström и др. 2020; Feuerborn и др. 2021). Арктическо-американская родословная включает в себя останки ездовых собак возрастом 9500 лет с острова Жохова в Сибири, несколько собак среднего голоцена с озера Байкал и других мест по всей Сибири, древних североамериканских собак, контактировавших друг с другом, и современные породы ездовых собак, такие как сибирский хаски и гренландская ездовая собака (Sinding и др. 2020;Feuerborn и др. 2021). Примечательно, что многочисленные останки собак и деревянных саней со стоянки Жохов в Арктической Сибири убедительно свидетельствуют о том, что собаки уже 9500 лет назад тянули сани (Pitulko and Kasparov 2017; Sinding и др. 2020). Эта филогенетическая группа современных и древних ездовых собак подтверждает генетическую преемственность и указывает на то, что ездовые собаки появились более 9500 лет назад на северо-востоке Сибирской Арктики (Sinding и др. 2020). Тем не менее, в то время как предыдущие исследования показали, что древняя американско-арктическая родословная доконтактных собак была в основном утрачена после колонизации Америки (Ní Leathlobhair и др., 2018), геномная история и наследие северо-восточной евразийской арктической родословной ездовых собак остаются в значительной степени неисследованными.
Археологические находки свидетельствуют о том, что ко времени появления собак Жохова ездовые собаки находились на продвинутой стадии одомашнивания. Питулько и Каспаров (2017) пришли к выводу, что на древней стоянке в Жохове присутствовали по крайней мере две полностью одомашненные и стандартизированные породы собак, причем 10 из 11 особей “полностью соответствовали современному стандарту сибирского хаски” по массе тела и росту, а вторая, более крупная порода была похожа по размерам на современного аляскинского маламута (далее - маламута) и гренландскую ездовую собаку. Вероятно, это не совпадение, поскольку мы ожидали бы тщательного отбора для достижения оптимальной эффективности у служебных собак. У ездовых собак, используемых для перевозки на дальние расстояния, более крупные собаки не могут достаточно быстро выделять тепло, а у маленьких собак возникают проблемы с сохранением тепла и поддержанием связи с упряжкой более крупных собак; таким образом, было доказано, что размер тела от 38 до 55 фунтов (от 17,2 до 24,9 кг) является оптимальным для собак, которые тянут упряжку на небольшие нагрузки на большие расстояния (Phillips и др., 1981). Морфометрические различия между этими двумя формами предполагают, что люди, возможно, уже отбирали собак для выполнения различных функций: фенотип, похожий на сибирского хаски, вероятно, лучше подходит для перевозки саней на большие расстояния, а более крупный - для перевозки грузов, охоты или защиты от крупных хищников. Более поздние находки арктической собаки возрастом 2000 лет, найденные на стоянке Ахим-Маяк на Западной Чукотке, также соответствуют современному стандарту породы сибирского хаски (Питулько и Каспаров, 2017) и могут свидетельствовать о временной неизменности размера и веса по крайней мере одной формы арктической собаки в этом регионе.
Ездовые собаки, обитавшие на крайнем северо-востоке Сибири, к востоку от реки Индигирки, и ассоциировавшиеся преимущественно с чукчами, коряками, юкагирами и камчадалами, были прародителями современной породы сибирского хаски (Томас и Thomas 2015). В прибрежных регионах эти ездовые собаки были жизненно необходимы для выживания людей, которые в основном добывали пропитание охотой на моржей и тюленей на морском льду (Богословская, 2022). Ездовые собаки также использовались для передвижения по горной тундре в суровых климатических условиях и с зимами, которые длятся большую часть года. Действительно, возможно, именно эта суровая и изолированная окружающая среда способствовала исторической генетической изоляции ездовых собак северо-восточной Сибири. Это контрастирует с популяциями северо-западных сибирских собак, которые, по-видимому, за последние несколько тысяч лет неоднократно подвергались смешению с представителями евразийских степей и Европы (Feuerborn и др., 2021). Хотя к концу 1700-х годов российская экспансия достигла крайнего северо-востока, а к началу 20-го века советская коллективизация привела к серьезным конфликтам в этом регионе (Кузина, 2022). Многие состоятельные фермеры, которые также разводили собак, исчезли (Крейн, 1977), что привело к потере знаний и навыков, связанных с оленеводством и ездой на собачьих упряжках (Кузина, 2022). Примерно в то же время большое количество сибирских собак было завезено на Аляску для участия в гонках на собачьих упряжках против гораздо более крупных местных аляскинских ездовых собак и других собак, завезенных на Аляску во время золотой лихорадки. Часть многих сибирских (на самом деле чукотских - мое дополнение, так как западные авторы постоянно путают русские географические названия) собак, которые прибыли на Аляску в период с 1908 по 1930 год или произошли непосредственно от собак, завезенных прямым импортом из Сибири на Аляску, были признаны Американским клубом собаководства (AKC) в 1930 году в качестве основы породы сибирского хаски.
К середине 20-го века в России катание на собачьих упряжках было признано экономически неэффективным и наносящим ущерб рыболовству (Кузина, 2022). Многие ездовые собаки были убиты или скрещены с иностранными разведенными породами, чтобы получить более крупных собак, которые могли бы тащить более тяжелые грузы, охотиться и приносить добычу (Хэндфорд, 1998; Кузина, 2022). Однако считается, что чукчи, скорее всего, содержали своих ездовых собак без примеси инородных организмов, особенно в изолированных деревнях вдоль северо-восточного побережья, где суровый климат также влияет на выживание (Томас и Томас 2015; Поцелуева 2022). К 1970 году популяция ездовых собак в Восточной Сибири сократилась из-за современного транспорта, сокращения охоты и рыболовства, нехватки продовольствия и болезней, завезенных иностранными собаками (Богословская, 2022). Российские кинологи не признали чукотскую ездовую собаку породой, что в сочетании с указом и усилиями по скрещиванию ее с иностранными собаками, не имеющими упряжки, может свидетельствовать о местном истреблении или замене генома за прошедшее столетие. В 1988 году обследование собак в районах и деревнях Чукотки показало, что осталось 1594 собаки (по сравнению с 50 700 в 1937 году только на Камчатке), но только около 400 из них, вероятно, были арктического происхождения, а остальные считались помесями с различной родословной ездовых собак (Богословская, 2022). Попытки восстановить породу начались в 1980-х годах, и в конце 20-го века Российская кинологическая федерация признала ее чукотской ездовой собакой (CSD).
Признание породы клубом собаководства в первую очередь служит двум целям: изоляции генофонда и ведению родословной. Запрещая скрещивание между признанными породами, клубы собаководства устанавливают искусственные барьеры для разведения, которые обеспечивают изоляцию генофонда и предоставляют подробную информацию о происхождении зарегистрированных собак. Следовательно, все генетическое разнообразие, которым когда-либо будет обладать порода, признанная клубом собаководства, заложено в генофонде при формировании породы, за исключением любых разрешенных дополнений в племенную книгу или недопустимого скрещивания и последующей фальсификации происхождения и родословной. Здесь мы исследуем влияние породных барьеров на генетическое здоровье сибирских хаски и проверяем популяционную дифференциацию внутри породы в зависимости от того, как собаки используются людьми (шоу, ездовые-шоу, гонки, домашние питомцы или собаки Сеппалы). Мы определяем выставочных собак как тех, кто участвует в выставках по конформации и, как правило, разводится в соответствии с письменными стандартами клуба собаководства. Ездовые-выставочные собаки также участвуют в соревнованиях по конформации, но также используются для катания на санях и других спортивных состязаний. Гоночных собак разводят для выполнения функциональных задач (например, для перевозки саней в туристических питомниках или для участия в гонках на собачьих упряжках) и могут быть выведены, а могут и нет, с определенным эстетическим видением. Домашних собак обычно разводят с единственной целью - продавать их населению (сюда не входят собаки из других популяций, которые также используются в качестве домашних питомцев). Наконец, собаки Сеппалы ведут свою историю от нескольких сибирских собак, которые были проданы в один питомник в 1930-х годах, и разделены на две породы. Недавно Континентальный клуб собаководства присвоил сибирской породе Сеппалы статус породы, хотя многие из них все еще зарегистрированы как сибирские хаски в других клубах собаководства по всему миру.
Чтобы оценить влияние закрытых генофондов и породных барьеров на генетическое здоровье, авторы использовали данные Зоономии о 240 млекопитающих (Консорциум Зоономии, 2020; Кристмас и др., 2023) и измерили уровни гомозиготности (ROH), чтобы выявить вредные варианты и оценить уровни генетического разнообразия, которые помогут сохранить эти эволюционно уникальные виды. родословные собак. Они также использовали сибирского хаски в качестве модели для оценки геномного наследия евразийской арктической линии, используя данные о однонуклеотидном полиморфизме (SNP) по всему геному 358 псовых, включая 280 зарегистрированных сибирских хаски и 5 современных CSD из России. Авторы проанализировали эти геномные данные, чтобы исследовать эволюционное родство, структуру популяции и западноевразийскую примесь. Они оценили общее происхождение современных ездовых собак и древних арктических псов и реконструировали глубокую популяционную историю арктических собак, используя калибровку ископаемых находок, чтобы оценить время расхождения.
Результаты
Структура популяции
Чтобы исследовать наследие северо-восточно-евразийской арктической линии современных собак, мы сначала провели геномное исследование, чтобы изучить уровни изоляции и примеси у сибирских хаски (далее - сибирский хаски). Анализ главных компонент (PCA) (рис. 1), ADMIXTURE (рис. 2) и филогенетическая реконструкция (рис. 3 а) выявляют значительную структуру популяции и предполагают, что искусственная изоляция внутри породы, основанная на том, как они используются людьми, привела к существенной генетической дифференциации в течение небольшого эволюционного периода (<100 лет). Эти популяции находятся в почти непрерывной зоне на PCA (рис. 1), где популяции шоу и Сеппалы образуют расходящиеся генетические кластеры. Результаты исследований (Alexander и др., 2009; Alexander и Lange, 2011) также свидетельствуют о том, что поток генов между современными популяциями сибирских хаски невелик, и в сибирской породе выделяются четыре генетических кластера: выставочный, домашний, Сеппала и гоночный (рис. 2). Помимо выставочной популяции ездовых собак, несколько сибирских хаски имеют небольшое количество предков из других сибирских популяций, но по большей части объединяются в свою соответствующую родословную, о которой сообщают их владельцы. Однако выставочно-ездовые особи не группировались ни одним из этих методов, а вместо этого группировались с выставочными, домашними, гоночными собаками или смесью всех других сибирских популяций.
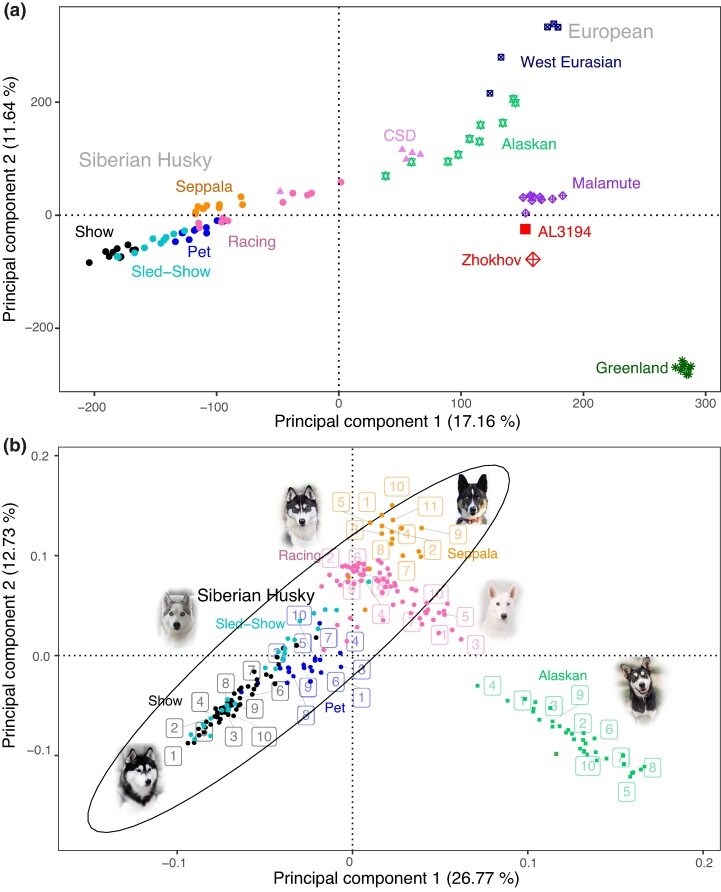
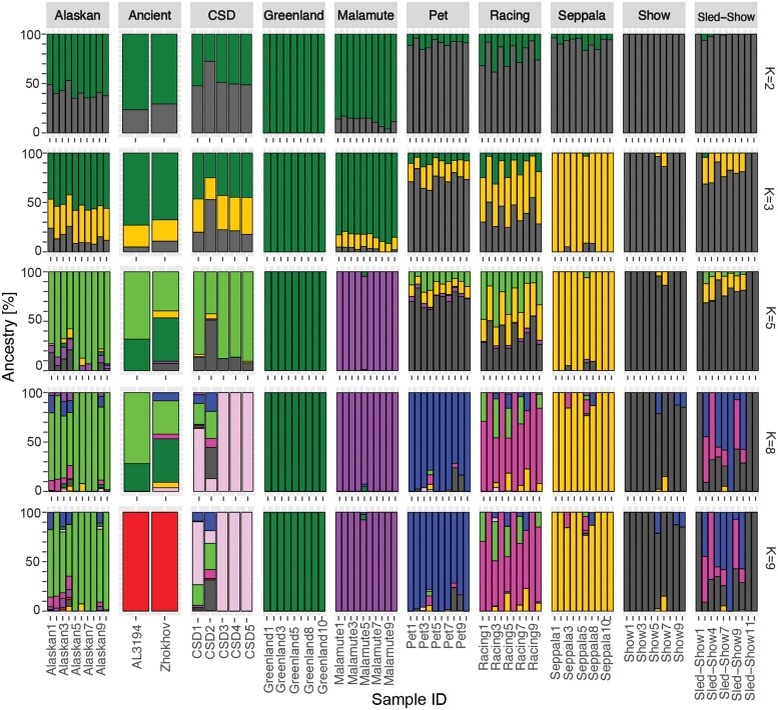
Чтобы проверить наличие сложной структуры популяции, авторы исследования определили количество оптимальных генетических кластеров с помощью k-средних, которые впоследствии были использованы в дискриминантном анализе главных компонент (DAPC). Популяции сибирских хаски, участвовавшие в шоу, соревнованиях на санях, домашних питомцев и гонках, были объединены в единый кластер, а популяция Сеппалы образовала уникальный кластер; однако половина представителей расы (расы 1, 3, 5, 7 и 10) и четыре представителя чукотских ездовых собак (CSD) были отнесены к генетическому кластеру Аляски/Западной Евразии. Однако, когда k был увеличен до 9, домашние, гоночные и выставочные сибирские хаски, а также западноевразийские собаки также сформировали свои собственные генетические кластеры. Кроме того, при k = 9 все сибирские хаски были также отнесены к популяции, о которой сообщил владелец, за исключением ездовых-шоу хаски, которые были объединены в группы выставок, гонок или домашних питомцев. Четыре CSD по-прежнему были связаны с аляскинскими ездовыми собаками, а один CSD - с популяцией хаски - домашних питомцев.
Расположение эллипса 99 %-ной достоверности вокруг породы Сибирский хаски с удаленными близкородственными особями показано на рисунке. на рис. 1б не включены 13 сибирских скакунов, найденных между группами сибирских и аляскинских ездовых собак. Генетический поток между аляскинскими ездовыми собаками (далее - аляскинцы) и некоторыми сибирскими скакунами также очевиден при анализе примесей (рис. 2). При K = 8 было обнаружено, что половина сибирских скакунов (5/10) имеют от 15 до 40% общего происхождения с аляскинцами, а трое из них также имеют небольшое количество общего происхождения с маламутами (<2%), что также характерно для современных аляскинцев (Huson и др., 2010). Четыре из этих собак (гонки 1, 3, 5 и 7) были обнаружены за пределами эллипса достоверности на PCA на рис. 1b, в то время как пятая (гонка 10) находилась прямо внутри эллипса, и все пять этих собак были объединены с аляскинскими/западноевразийскими собаками в DAPC (дополнительные фиги. Разделы 3 и 4, Дополнительные материалы в Интернете).
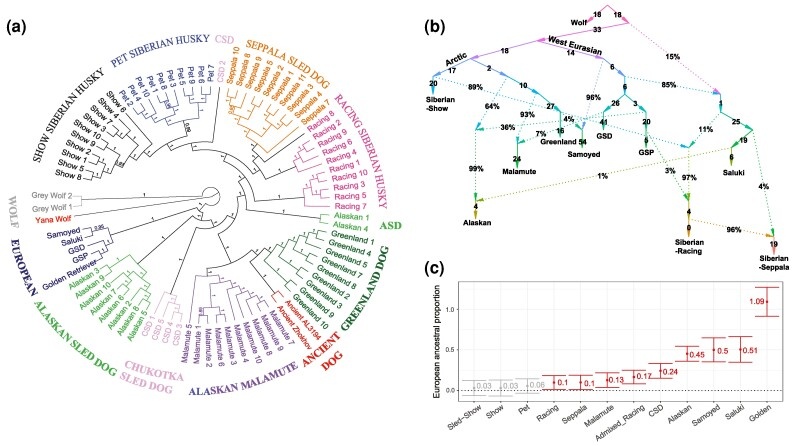
Филогенетическая реконструкция, в соответствии с результатами, полученными методом ADMIXTURE , также выявила четыре генетически отличающиеся популяции в породе: выставочная, домашняя, Сеппала и гоночная (рис. 3 а). Однако, в то время как выставочные популяции, популяции домашних питомцев и Сеппалы были монофилетическими, было обнаружено, что гоночная популяция хаски была парафилетической с пятью особями, которые были идентифицированы как имеющие аляскинское происхождение в результате анализа примесей (рис. 2), которые также были объединены с аляскинскими и западноевразийскими собаками в DAPC, и были обнаружены за пределами 99 %-ного доверительного интервала по шкале PCA (рис. 1 b), как группа, отличающаяся от остальных представителей породы. Эти собаки также сформировали свою собственную группу, несмотря на значительное географическое разделение между этими особями (трое из Северной Америки, одна из Европы и одна из Океании) и отсутствие недавнего общего родства (коэффициент родства < 0,088).
Западноевразийская примесь в арктической родословной
Чтобы лучше понять сложную эволюционную историю породы сибирского хаски и то, как она в более широком смысле соотносится с историей других арктических и древних собак, авторы реконструировали историю популяции, используя qpGraph (рис. 3 b) и qpAdm, как это реализовано в ADMIXTOOLS v2.0.0 (Patterson и др. 2012; Maier и др. 2023). Порода была выведена с учетом существенного различия между популяциями, что согласуется с предыдущими результатами авторов. Значительное европейское происхождение было смоделировано в популяциях гоночных собак и Сеппала (рис. 3 b). Метод QpAdm, который проверяет наличие вероятных популяций-предков, также подтвердил модель значительной европейской примеси в этих популяциях (P > 0,05) и вывел аналогичные пропорции примеси. Напротив, выставочные и ездовые популяции сибирских хаски и гренландских ездовых собак показали отсутствие существенной примеси какой-либо из включенных или родственных пород, а также не включенных в выборку линий-призраков.
Затем, чтобы проверить наличие генов у западноевразийских пород, которые, как было установлено ранее, присутствуют в геноме современных аляскинских ездовых собак (Huson и др., 2010; Thorsrud и Huson, 2021), были протестированы все собаки из исследуемого набора данных на интрогрессию из Европы, используя D-статистику Паттерсона (рис. 4), статистика f3 и коэффициента f4 (рис. 3 с). В ходе анализа авторы обнаружили значительную интрогрессию с несколькими западноевразийскими породами у всех аляскинских собак, кроме одной (рис. 4 а). В тесте на соотношение f4 также было показано, что аляскинские ездовые собаки имеют ~45 % европейского происхождения (|Z| = 14,811) (рис. 3 с). Таким образом, если бы генетический поток шел от аляскинцев к сибирским гоночных хаски, как предполагает анализ ADMIXTURE , были бы обнаружена значительная интрогрессия, по крайней мере, с некоторыми западноевразийскими породами, у сибирских хаски, имеющих общих предков с аляскинцами.
Было обнаружено, что около 17 % пород сибирских гоночных хаски имеют европейское происхождение (|Z| = 6,041), а также значительную интрогрессию с немецким короткошерстным пойнтером (GSP), немецкой овчаркой (GSD), салюки и золотым ретривером (|Z| ≥ 3,925), которые находятся за пределами эллипса достоверности (рис. 3 с), что согласуется с анализами Qpgraph и QPADM. Эти сибирские хаски также имеют значительно больше наследственных аллелей с европейскими породами по сравнению со всеми другими популяциями. Кроме того, все собаки, имеющие общее происхождение с аляскинскими ездовыми собаками в анализе ADMIXTURE (Alexander и др., 2009; Alexander и Lange, 2011) на рис. 2 (K = 8), также имеют значительную интрогрессию по крайней мере с одной из европейских пород, включенных в этот анализ (рис. 4 а), что свидетельствует том, что у этих собак происходит переход генов от аляскинских или западноевразийских к сибирским линиям. В дальнейшем здесь эти сибирские хаски-гонщики будут называться “смешанной расовой группой”.
Аналогичным образом, было обнаружено, что у сибирских гоночных собак и Сеппала наблюдается значительная интрогрессия по GSP (|Z| > 3,261) и салюки (|Z| > 3,566), а также тесты на соотношение f4 показали, что в обеих популяциях европейское происхождение составляет около 10 % (рис. 3 с), за исключением сибирских пород с примесью, которые находятся за пределами эллипса достоверности в PCA, что также согласуется с моделью примеси и пропорции, вычисленные с помощью qpgraph (рис. 3 b) и QPADM. На индивидуальном уровне было обнаружено, что около половины из 111 представителей гоночной и 24 представителей линии Сеппала, включенных в этот анализ, имеют значительную европейскую интрогрессию (рис. 4 b). Напротив, только 5 из 73 выставочных линий и 4 из 50 сибирских ездовых хаски имеют значительную европейскую интрогрессию (рис. 4), а также ни в одной из аляскинских популяций не было обнаружено существенного вклада примесей от европейских собак (рис. 2) по данным ADMIXTURE, коэффициента f4 (рис. 3 c), D-статистики, qpGraph (рис. 3 b и 5 b), и qpAdm. Подобно современной аляскинской, чукотские ездовые собаки испытали на себе сопоставимое влияние европейских пород у всех пяти современных образцов, включенных в этот анализ, особенно со стороны GSD (рис. 4 a, 5 b и c).
Древний арктический предок
Было установлено, что гренландская ездовая собака является ближайшим из ныне живущих родственников доконтактных собак, включенных в этот анализ собак с острова Жохова и Порт-о-Шуа до контакта с человеком (рис. 3 а, 5 b и c). Анализ примесей продемонстрировал близкое родство 4000-летней собаки породы Порт-о-Шуа (AL3194) с американскими ездовыми собаками, причем большинство (72 %) ее предков относятся к аляскинцам, а 28 % - к гренландской ездовой собаке (K = 8; рис. 2). Собака Жохова также в основном была связана с американскими ездовыми собаками (34 % - с аляскинскими и 44 % - с гренландскими), но 22 % ее предков были связаны с сибирскими хаски и чукотскими ездовыми (K = 8; рис. 2). Было обнаружено, что все современные арктические собаки имеют значительную долю общего происхождения с линиями жоховских/американских доконтактных собак, но гренландская ездовая собака и маламут родственным с ними гораздо в большей степени (от 70 % до 72 %), нежели сибирские хаски (от 28 % до 42 %). Что касается сибирских собак, то линия Сеппала и современные чукотские ездовые имеют меньше всего общего с древней ездовой собакой Жохова (28 %), а выставочная популяция и популяции ездовых хаски, участвующие в выставках, имеют наибольшее количество общего происхождения (42 %). У похожего на современных европейских собак самоед, который, как было установлено, имеет в основном западноевразийское происхождение (рис. 3 а-с), не было обнаружено существенного родства с древними ездовыми собаками, согласно данным по коэффициенту f4, но у собаки Жохова имеется избыток производных аллелей по сравнению с европейскими собаками, что показывает D-статистика.
Глубокая популяционная история
Чтобы оценить время расхождения в пределах арктической клады, авторы сначала использовали модель Fossilized birth–death (FBD) model (Heath и др., 2014) и Optimised Relaxed Clock (ORC) (Douglas и др., 2021) в байесовском филогенетическом анализе BEAST2 (Bouckaert и др., 2014, 2019). Этот анализ полностью поддерживает топологию, соответствующую полученным данным
с помощью Treemix (Pickrell и Pritchard 2012) и FastTree (как это реализовано в GENEIOUS PRIME v2023.0.4; https://www.geneious.com) результаты (рис. 3 а и 5 с), что указывает на присутствие, по крайней мере, двух линий арктических собак на северо-востоке Арктической Сибири в конце плейстоцена/начале голоцена (95 %-ная наибольшая плотность потомства [HPD]: от 9700 до 14 300 лет назад). Оценки времени расхождения арктических линий с помощью SNAPP (Bryant и др., 2012) и методов Stange и др. (2018) также подтвердили эти результаты (рис. 5 а), установив расхождение между этими двумя современными арктическими линиями в конце плейстоцена - около 12800 лет назад.
Далее, чтобы исследовать сложную популяционную историю этих различных линий арктических собак, авторы исследовали древнюю примесь, используя функцию find_graph() в qpGraph (рис. 5 b) (Patterson и др., 2012; Maier eи др., 2023). Наиболее подходящая модель f-статистики показывает, что собака Жохова имеет большую часть своей родословной с арктическими собаками, но около 36 % происходит от древних волков. Исследование Treemix (Pickrell и Pritchard, 2012) также показало примесь плейстоценовых волков у жоховских и гренландских ездовых собак, а также интрогрессию европейских пород у маламутов, аляскинских и чукотских ездовых собак (рис. 5 с). Напротив, никакой интрогрессии со стороны волков не было обнаружено у большого числа современных сибирских хаски и аляскинских ездовых собак, а также у нескольких аляскинских маламутов и CSD, включенных в это исследование. Это согласуется с данными D-статистики, которые показали значительную плейстоценовую интрогрессию волков у Жохова, контактной собаки Порт-о-Шуа AL3194 и гренландских ездовых собак, но ее отсутствие у других арктических собак.
Наконец, чтобы исследовать фенотипическое сходство с современными породами ездовых собак, была создана реконструкция того, как выглядела древняя собака Жохова, исходя из последовательности генома. В сочетании с данными ASIP, полученными от Bannasch и др. (2021), и данными IGF1-AS, полученными от Plassais и др. (2022), собака Жохова, которая была обнаружена на первом участке плейстоценовой стоянки и представлена второй по величине нижней челюстью, вероятно, была среднего размера, с двойной шерстью, черного цвета с подпалинами очки и умеренное количество белых пятен, карими глазами (рис. 5 d). Эта древняя собака была носителем рецессивного красного гена MC1R (e1) и аллеля, снижающего уровень феомеланина в гене MFSD12, который часто встречается у современных ездовых собак. Интересно, что у этой собаки не было ни голубых глаз, ни древнего гена (eA) рыжего окраса, что встречается у современных сибирских хаски, но отсутствует у гренландских ездовых собак.
Генетическое здоровье
Численность популяции
Как и у многих современных пород собак, у современных арктических пород эффективная численность популяции невелика - от 41 особей у маламутов до 192 особей у гренландских ездовых собак и 571 особей у домашних сибирских хаски (рис. 6 а). Среди сибирских хаски в популяциях - выставочной и Сеппала - наблюдается наибольший уровень неравновесия по сцеплению (LD) и наименьший эффективный размер популяции (Ne = 79 и 47 соответственно). В то время как у сибирских хаски и гренландских ездовых собак численность популяции превышает 50 особей, у сибирских маламутов и линии Сеппала - нет. У всех протестированных здесь пород, за исключением популяции домашних хаски, показатель Ne < 200, по сравнению с 3000 у предков сибирского хаски и аляскинской ездовой собаки и 500 у гренландской ездовой собаки всего 125 поколений назад (рис. 6 а).
Инбридинг
Используя породу сибирский хаски и ее отдельные популяции с различным уровнем примеси в качестве модели для исследования влияния закрытых генофондов на генетическое разнообразие, авторы обнаружили, что относительно несмешанная выставочная популяция имела самую высокую среднюю долю генома, содержащуюся в ROH (FROH = 31 %) (рис. 6 b) и самую низкую гетерозиготность среди всех сибирских популяций. В отличие от этого, у расовой популяции средний показатель FROH был значительно ниже - 18 % (критерий Вилкоксона, P = 2,2*10-16) и наблюдалась самая высокая гетерозиготность из всех сибирских популяций. Внутри породы доля пар собак в каждой популяции, имеющих общие сегменты ДНК, идентичные по происхождению (IBD), колеблется от 31 % в популяции гоночных собак до 90 % в популяции Сеппалы. У гренландской ездовой собаки также очень высокая доля пар собак с общими сегментами IBD (92 %), в отличие от аляскинской ездовой собаки, у которой она составила только 7 %.
Потенциально опасные варианты
Используя привязанную к собакам версию таблицы Zoonomia "240 видов млекопитающих кактусов" для обозначения позиций, которые были эволюционно ограничены у сибирских хаски (Zoonomia Consortium, 2020; Christmas и др., 2023), было обнаружено, что популяции Сеппалы и домашних сибирских хаски имеют значительно более высокую частоту мутировавших эволюционно ограниченных вариантов по сравнению с другими популяциями, участвующими в гонках (рис. 6 c). Кроме того, популяция Cеппалы также имела значительно больше вариантов, которые, по прогнозам, были вредными, используя аннотацию SnpEff (Cingolani и др., 2012) (рис. 6 d).
Обсуждение
Популяционаая структура
Структура популяции является следствием неслучайного скрещивания, когда изолированные популяции имеют меньше общих предков, чем можно было бы ожидать случайно. Когда собак используют для разных целей, они становятся более специализированными, приобретая черты, которые способствуют достижению этой цели. Это приводит к усилению изоляции, поскольку люди склонны искусственно отбирать особей внутри популяции из-за предполагаемой потери качества породы при скрещивании ее особей с особями популяций, которые не были выведены для их конкретной цели. Поэтому неудивительно, что структура популяции породы сибирский хаски основана на том, как они используются людьми, и это расхождение между популяциями будет продолжать увеличиваться с течением времени, если не произойдет существенного обмена генами между популяциями.
Скрещивание аляскинских ездовых собак или европейских пород с сибирскими хаски, возможно, было использовано для получения аллелей, повышающих производительность, от более быстрых и жаростойких пород, что было бы выгодно, поскольку гонки на собачьих упряжках распространились и в более теплом климате. Аляскинская ездовая собака, выведенная в первую очередь для спортивных состязаний и не получившая официального признания кинологических клубов, стала ведущим участником гонок на собачьих упряжках, включая Идитарод, выиграв множество соревнований на различных дистанциях. Хотя изначально они происходят от местных деревенских собак на Аляске, сегодня их в основном специально разводят для спринтерских гонок и гонок на длинные дистанции, и в их генетический состав вошло много различных пород в результате скрещивания с европейскими породными группами, такими как борзые и пойнтеры, которые использовались на Аляске в течение последнего столетия для улучшения скорости и жаростойкости (Huson и др., 2012). Они выработали свой собственный уникальный геномный признак породы (Huson и др., 2010; Торсруд и Хусон, 2021), что подтверждается приведенными здесь данными, тем не менее, они по-прежнему имеют значительную долю общего происхождения с древними ездовыми собаками.
Было также обнаружено, что современные чукотские ездовые имеют значительную примесь европейских пород собак (рис. 3 с, 4 и 5 b, c) на арктическом генетическом фоне, что может объяснить их близость к аляскинцам по PCA и DAPC (рис. 1). Систематическое разведение ездовых собак в России для создания более крупной грузовой собаки (Хэндфорд, 1998), а также советская коллективизация и миграция людей в Восточную Сибирь вместе с западными собаками (Богословская, 2022), вероятно, являются объяснением значительной интрогрессии этих современных европейских пород собак в аборигенные популяции собак Сибири. Кроме того, две из взятых в исследование чукотских ездовых собак в значительной степени имеют общее происхождение с современными сибирскими хаски и аляскинцами (рис. 2), а у одной также есть общие сегменты IBD с большим количеством современных выставочных и домашних сибирских хаски, что указывает на недавнюю примесь с сибирскими хаски. Эти данные свидетельствуют о том, что современные сибирские хаски, за исключением примерно половины тех, кто участвует в гонках, могут быть обладателями наименее смешанной евразийско-арктической родословной, оставшейся без существенной недавней западно-евразийской примеси. Однако следует отметить, что было отобрано только пять особей из двух разных питомников. Дополнительная выборка может выявить более изолированные популяции с меньшим количеством западноевразийских примесей.
Древний арктический предок
Предыдущие исследования продемонстрировали непрерывную арктическую родословную собак на протяжении по меньшей мере 9500 лет (Brown и др., 2013; Sinding и др., 2020; Feuerborn и др., 2021). Также было обнаружено, что 9500-летняя ездовая собака с острова Жохова в Восточной Сибири имеет значительно больше общих генетических аллелей с современными арктическими собаками по сравнению с современными европейскими собаками (рис. 5 b), в которых обобщены полученные филогенетические данные (рис. 3 a и 5 c), демонстрирующие тесное филогенетическое родство между древними и современными ездовыми собаками. Как и Фейерборн и др. (2021), авторы также обнаружили, что гренландская ездовая собака является близкой родственницей древних сибирских и американских ездовых собак (рис. 3 a и 5 c). В соответствии с этой филогенетической топологией было отмечено, что гренландская ездовая собака ведет большую часть своей родословной от этой древней линии, в то время как современные сибирские хаски имеют гораздо меньшее родство с нею (рис. 2 и 5 b). Недавнее исследование показало, что это, вероятно, было связано с примесью у породы сибирского хаски, и продемонстрировало значительную европейскую интрогрессию у не-гренландских пород ездовых собак, таким образом, сделав вывод, что гренландская ездовая собака имеет больше всего общего генома с линией Жохова, потому что они наименее "загрязнены" примесями (Sinding и др., 2020). Однако было обнаружено, что европейская примесь в основном присутствует в той части сибирских хаски, которые используются для гонок, а другие сибирские популяции в последнее время не претерпели значительной западноевразийской интрогрессии. Более того, авторами показано, что маламут имеет значительную примесь европейских пород (от 13 % до 16 %, Z ≥ 4,248 ; рис. 3 с) и ведет свою родословную от собаки Жохова в большей степени, нежели относительно не смешанная ни с кем выставочная популяция сибирских хаски (70 % против 42 %). Таким образом, недавняя западноевразийская примесь, вероятно, не является достаточным объяснением того, почему гренландская ездовая собака и маламут имеют больше общего происхождения с древней собакой Жохова, чем арктические собаки из Сибири. Таким образом, мы считаем, что большее генетическое сходство между гренландскими собаками и древней собакой Жохова, по сравнению с сибирскими хаски, отражает не уровень примесей в современных сибирских собаках, а порядок, в котором эти линии собак отделились друг от друга, что подтверждается их филогенетическим расположением.
Глубокая популяционная история
Чтобы проверить гипотезу о древней структуре популяции, авторы исследовали временные расхождения в арктической линии, используя древних собак, чтобы соотнести относительное время с оценкой абсолютного возраста. Важно отметить, что они провели масштабный анализ примесей в нашем своем данных об арктических собаках и включили в свое исследование сроков дивергенции только тех собак и породы, у которых не было недавней западноевразийской интрогрессии. Смешение между кладами затрудняет филогеномный анализ, приводя к завышению или недооценке возраста внутренних узлов, и, как было показано, является относительно распространенным явлением у современных породистых собак (Meadows и дрl., 2023), что также продемонстрировано в этом исследовании.
Полученные результаты свидетельствуют о древнем расколе между сибирскими и гренландско-жоховскими линиями ездовых собак в конце плейстоцена (рис. 5 а), что совпадает с резкими климатическими сдвигами, происходящими при переходе Земли от ледникового периода к более теплому и влажному межледниковому циклу, характеризующемуся потерей местообитаний и последующим массовым вымиранием мегафауны (Monteath и др., 2021). Это, возможно, положило начало или ускорило использование псовых для транспортировки на большие расстояния, а также для охоты и транспортировки более мелкой дичи, такой как олени карибу и различные морские млекопитающие, что привело к селекции собак для различных целей. Гипотеза о более раннем отделении предков сибирского хаски от жоховской/гренландской линии также подтверждается общим дрейфом арктического и американского геномов, который наблюдался от Гренландии через Берингов пролив через Сибирь в Европу. Кроме того, Жоховский и Гренландский расколы произошли непосредственно перед или во время затопления Берингова перешейка, что затруднило и в конечном итоге предотвратило миграцию животных примерно с период 11000-9500 лет назад (Элиас и др., 1996) и, вероятно, представляет собой более позднее расселение собак по Северной и Южной Америке после первоначального раскола, обнаруженного в предыдущих исследованиях (Ní Leathlobhair и др., 2018; Ameen и др., 2019). Филогенетический анализ также позволяет предположить, что современные гренландские собаки происходят от линии, которая совсем недавно отделилась от доконтактной собаки Порт-о-Шуа, по сравнению со всеми другими современными арктическими линиями, что подтверждается избытком общих наследственных аллелей и большим общим генетическим дрейфом с гренландскими собаками в сравнении с любой другой современной арктической породой.
Предки плейстоценового сибирского волка ранее были обнаружены в родословной арктической собаки (Skoglund и др., 2015; Ní Leathlobhair и др., 2018; Синдинг и др., 2020; Фейерборн и др., 2021; Рамос Мадригал и др., 2021). Здесь, однако, эта древняя волчья примесь обнаруживается и у гренландских, жоховской и доконтактной (AL3194) собак, однако доказательств избыточного распределения аллелей у большого количества современных сибирских хаски или других ездовых собак по сравнению со всеми другими протестированными арктическими и европейскими породами не было найдено, и ни сибирские хаски, ни чукотские ездовые не вписывались в модель с примесью плейстоценовых волков Сибири, с использованием qpGraph и Treemix (рис. 5 b и c). Предыдущие исследования также показали наличие явных признаков присутствия янского волка у гренландских и жоховских собак, но свидетельства древней интрогрессии волков у сибирских хаски были гораздо слабее (Sinding и др., 2020; Feuerborn и др., 2021). В сочетании с полученными здесь результатами, можно сделать вывод, что наиболее логичным объяснением является то, что плейстоценовая популяция волков произошла после того, как сибирская родословная отделилась от жоховской/гренландской. Вполне возможно, что поток генов от плейстоценовых волков мог быть использован древними жителями Северной Сибири для увеличения размеров и охотничьих способностей предков жоховской/гренландской линии, что, возможно, было полезно, поскольку климат потеплел, животные мегафауны вымерли, а традиционные источники пищи, вероятно, стали дефицитными, но при этом сохранялись линии ездовых собак для перевозки и других видов деятельности, которые собаки выполняют лучше, чем гибриды с волками. Авторы не считают, что это открытие может быть объяснено различными пропорциями между восточными и западными предками-волками в кладе арктических собак, поскольку гренландские ездовые собаки и сибирские хаски имели столь же низкие пропорции с западными волчьими предками (Бергстрем и др., 2022), и ни одна из арктических собак в приведенном наборе данных в отличие от гренландских собак и собаки Жохова не проявляет сходство с плейстоценовыми волками даже по сравнению с современными западноевразийскими собаками. В качестве альтернативы, также возможно, что эта закономерность может быть результатом неучтенной примеси у сибирских хаски. Важно отметить, что это древнее смешение с волками в гренландско-жоховской линии также может внести путаницу в данные оценки времени расхождения. Дополнительные анализы, включающие более древние последовательности генома сибирских хаски и волка, помогут в будущем уточнить эти оценки времени расхождения.
Генетическое здоровье
Численность популяции
Среди сибирских собак выставочные популяции и популяция Сеппала имеют наибольшее количество LD и наименьший эффективный размер популяции (Ne = 79 и 47 соответственно), что, вероятно, связано с большим количеством инбридинга в этих популяциях, что приводит к устойчивому увеличению LD и сокращению эффективного размера популяции (рис. 6 а). Недавнее увеличение числа домашних сибирских хаски за последние десять поколений может быть отчасти связано с возросшей популярностью этой породы среди владельцев домашних животных и заводчиков-любителей; однако форма кривой также указывает на наличие недавней примеси, и поэтому эти данные следует интерпретировать с осторожностью. С точки зрения сохранения и жизнеспособных популяций, авторы стремятся к минимальному эффективному размеру популяции в 50 особей в краткосрочной перспективе, чтобы уменьшить потерю гетерозиготности и депрессию инбридинга, и к долгосрочному Ne > 500 для поддержания эволюционного потенциала и уменьшения случайных эффектов генетического дрейфа. Эти цифры были основаны на наблюдениях за домашними животными, которые продемонстрировали, что отбор по продуктивности и плодовитости не может преодолеть инбридинговую депрессию, если коэффициент инбридинга увеличивается более чем на 1 % за поколение (Franklin, 1980).
Инбридинг
Коэффициенты инбридинга, основанные на родословной (т.е. COI), дают оценку ожидаемой доли генома, который является IBD и содержится в протяженных участках гомозиготности (ROH). Используя общегеномные ДНК-маркеры, можно не только определить долю генома, которая находится в ROH, но и использовать длину ROH, чтобы определить, имели ли две линии общего предка с более длинными сегментами в результате более позднего инбридинга. Гоночная популяция имеет самый низкий уровень инбридинга из всех сибирских популяций, в то время как выставочная популяция имеет самый высокий уровень инбридинга (рис. 6 b). Важно отметить, что около половины случаев инбридинга у современных сибирских хаски происходят из-за недавнего инбридинга в течение последних ∼6 поколений. В отличие от этого, большинство случаев инбридинга у гренландских ездовых собак гораздо старше. Количество ROH у отдельных особей в зависимости от размера ROH указывает на то, что популяция гренландских ездовых собак в прошлом испытывала трудности по типу бутылочного горлышка, объединившего большое количество мелких фрагментов.
Большая доля геномов, содержащихся в ROH, в этих популяциях также подтверждается большим количеством общих IBD у пар особей в популяциях сибирских хаски, маламутов и гренландских ездовых собак. Почти каждая пара собак в популяции Сеппалы и гренландской популяции имеет часть своего генома, происходящую от общего предка, что затрудняет возможность спаривания родственников и увеличивает риск инбридинговой депрессии. Около двух третей пар собак, участвующих в выставках, выставках ездовых животных и домашних питомцах, имеют общие сегменты IBD, указывающие по крайней мере на одного недавнего общего предка. Напротив, только 31 % пар собак в популяции, участвующей в гонках, имеют общие сегменты IBD, что может быть объяснено недавним добавлением в эту популяцию свежей родословной, которое уменьшает распределение IBD (Blondeau Da Silva и др., 2024).
Потенциально опасные варианты
Мутации в позициях, которые эволюционно ограничены у разных видов, обогащаются болезнетворными вариантами (Sullivan и др., 2023). Аналогичным образом, мутации, которые приводят к потере функции белка, могут возникать у нездоровых фенотипов. Таким образом, частота этих типов вариантов в популяции является информативной для генетического здоровья. В гоночных, выставочных и ездовых-шоу популяциях сибирских хаски меньше потенциально опасных вариантов (рис. 2, 6 c и d), что может свидетельствовать о мощном селективном отборе по форме и функциям при сохранении эффективной численности популяции > 50, и что играет определенную роль в очистке геномов от некоторых вредных вариантов и снижении генетической нагрузки. С другой стороны, ослабление давления отбора в популяции домашних животных может привести к сохранению вариантов, которые без вмешательства человека или медицинской помощи обычно снижают работоспособность. Ездовые собаки линии Сеппала, вероятно, представляют собой пример критически низкой численности популяции (Ne = 47), что делает их восприимчивыми к случайным эффектам генетического дрейфа и снижает эффективность отбора. Несмотря на то, генетически линия Сеппалы отличается от других сибирских хаски, исходя из этих результатов, дальнейшая изоляция при разведении может быть вредной для здоровья популяции.
Выводы
Европейская колонизация, советская экспансия и преднамеренное скрещивание привели к интрогрессии западноевразийских пород в современные популяциях арктических собак, что потенциально угрожает геномной идентичности или исчезновением уникальных линий собак, которые существовали тысячи лет (Handford, 1998; Ní Leathlobhair и др., 2018; Sinding и др., 2020; Feuerborn и др. 2021; Богословская 2022). Здесь было обнаружено, что порода сибирский хаски (за исключением примерно половины собак, участвующих в гонках на собачьих упряжках) и гренландские ездовые собаки представляют собой две наименее смешанные популяции современных арктических собак, происходящие от древних линий собак с непрерывной историей, начиная с позднего плейстоцена. Однако современные породные барьеры и изоляция небольших популяций, ориентированных на конкретные цели, привели к значительной дифференциации и умеренному уровню инбридинга сибирских хаски в течение короткого периода эволюции, что создает проблемы для долгосрочного сохранения этой линии. Поскольку борьба за сохранение различных эволюционных линий при сохранении генетического здоровья породистых собак усиливается, понимание геномной истории имеет важное значение для разработки эффективной политики и наилучших методов управления породами.
Список литературы
- Alexander DH, Lange K. Enhancements to the ADMIXTURE algorithm for individual ancestry estimation. BMC Bioinfor. 2011:12(1):246. 10.1186/1471-2105-12-246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alexander DH, Novembre J, Lange K. Fast model-based estimation of ancestry in unrelated individuals. Genome Res. 2009:19(9):1655–1664. 10.1101/gr.094052.109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ameen C, Feuerborn TR, Brown SK, Linderholm A, Hulme-Beaman A, Lebrasseur O, Sinding MHS, Lounsberry ZT, Lin AT, Appelt M, et al. Specialized sledge dogs accompanied Inuit dispersal across the North American Arctic. Proc Biol Sci. 2019:286(1916):20191929. 10.1098/rspb.2019.1929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bannasch DL, Kaelin CB, Letko A, Loechel R, Hug P, Jagannathan V, Henkel J, Roosje P, Hytönen MK, Lohi H, et al. Dog colour patterns explained by modular promoters of ancient canid origin. Nat Ecol Evol. 2021:5(10):1415–1423. 10.1038/s41559-021-01524-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bergström A, Frantz L, Schmidt R, Ersmark E, Lebrasseur O, Girdland-Flink L, Lin AT, Storå J, Sjögren KG, Anthony D, et al. Origins and genetic legacy of prehistoric dogs. Science. 2020:370(6516):557–564. 10.1126/science.aba9572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bergström A, Stanton DW, Taron UH, Frantz L, Sinding MHS, Ersmark E, Pfrengle S, Cassatt-Johnstone M, Lebrasseur O, Girdland-Flink L, et al. Grey wolf genomic history reveals a dual ancestry of dogs. Nature. 2022:607(7918):313–320. 10.1038/s41586-022-04824-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blondeau Da Silva S, Mwacharo JM, Li M, Ahbara A, Muchadeyi FC, Dzomba EF, Lenstra JA, Da Silva A. IBD sharing patterns as intra-breed admixture indicators in small ruminants. Heredity. 2024:132(1):30–42. 10.1038/s41437-023-00658-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bogoslovskaya L. 2022. Sled dogs of Russia. In: Beregovoy V, editor. The dogs of our ancestors . India: CATNUS Publishers. p. 459–462. [Google Scholar]
- Botigué LR, Song S, Scheu A, Gopalan S, Pendleton AL, Oetjens M, Taravella AM, Seregély T, Zeeb-Lanz A, Arbogast RM, et al. Ancient European dog genomes reveal continuity since the Early Neolithic. Nat Commun. 2017:8(1):16082. 10.1038/ncomms16082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouckaert R, Heled J, Kühnert D, Vaughan T, Wu CH, Xie D, Suchard MA, Rambaut A, Drummond AJ. BEAST 2: a software platform for Bayesian evolutionary analysis. PLoS Comput Biol. 2014:10(4):e1003537. 10.1371/journal.pcbi.1003537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouckaert R, Vaughan TG, Barido-Sottani J, Duchêne S, Fourment M, Gavryushkina A, Heled J, Jones G, Kühnert D, De Maio N, et al. BEAST 2.5: an advanced software platform for Bayesian evolutionary analysis. PLoS Comput Biol. 2019:15(4):e1006650. 10.1371/journal.pcbi.1006650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown SK, Darwent CM, Sacks BN. Ancient DNA evidence for genetic continuity in Arctic dogs. J Archaeol Sci. 2013:40(2):1279–1288. 10.1016/j.jas.2012.09.010. [DOI] [Google Scholar]
- Bryant D, Bouckaert RR, Felsenstein J, Rosenberg NA, RoyChoudhury A. Inferring species trees directly from biallelic genetic markers: bypassing gene trees in a full coalescent analysis. Mol Biol Evol. 2012:29(8):1917–1932. 10.1093/molbev/mss086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Christmas MJ, Kaplow IM, Genereux DP, Dong MX, Hughes GM, Li X, Sullivan PF, Hindle AG, Andrews G, Armstrong JC, et al. Evolutionary constraint and innovation across hundreds of placental mammals. Science. 2023:380(6643):eabn3943. 10.1126/science.abn3943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cingolani P, Platts A, Wang LL, Coon M, Nguyen T, Wang L, Land SJ, Lu X, Ruden DM. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012:6(2):80–92. 10.4161/fly.19695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crane B. The rise and decline of Russia's Sled dogs. In: International Siberian Husky Club presents the Siberian Husky. Oregon: Lee's Print Shop; 1977. p. 25–26. [Google Scholar]
- Douglas J, Zhang R, Bouckaert R. Adaptive dating and fast proposals: revisiting the phylogenetic relaxed clock model. PLoS Comput Biol. 2021:17(2):e1008322. 10.1371/journal.pcbi.1008322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feuerborn TR, Carmagnini A, Losey RJ, Nomokonova T, Askeyev A, Askeyev I, Askeyev O, Antipina EE, Appelt M, Bachura OP, et al. Modern Siberian dog ancestry was shaped by several thousand years of Eurasian-wide trade and human dispersal. Proc Natl Acad Sci U S A. 2021:118(39):e2100338118. 10.1073/pnas.2100338118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Franklin IA. Evolutionary change in small population. Sunderland: Sinauer Associates; 1980. [Google Scholar]
- Frantz LA, Mullin VE, Pionnier-Capitan M, Lebrasseur O, Ollivier M, Perri A, Linderholm A, Mattiangeli V, Teasdale MD, Dimopoulos EA, et al. Genomic and archaeological evidence suggest a dual origin of domestic dogs. Science. 2016:352(6290):1228–1231. 10.1126/science.aaf3161. [DOI] [PubMed] [Google Scholar]
- Freedman AH, Gronau I, Schweizer RM, Ortega-Del Vecchyo D, Han E, Silva PM, Galaverni M, Fan Z, Marx P, Lorente-Galdos B, et al. Genome sequencing highlights the dynamic early history of dogs. PLoS Genet. 2014:10(1):e1004016. 10.1371/journal.pgen.1004016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Handford JM. Dog sledging in the eighteenth century: North America and Siberia. Polar Rec. 1998:34(190):237–248. 10.1017/S0032247400025705. [DOI] [Google Scholar]
- Heath TA, Huelsenbeck JP, Stadler T. The fossilized birth–death process for coherent calibration of divergence-time estimates. Proc Natl Acad Sci U S A. 2014:111(29):E2957–E2966. 10.1073/pnas.1319091111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huson HJ, Parker HG, Runstadler J, Ostrander EA. A genetic dissection of breed composition and performance enhancement in the Alaskan sled dog. BMC Genet. 2010:11(1):71–14. 10.1186/1471-2156-11-71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huson HJ, Vonholdt BM, Rimbault M, Byers AM, Runstadler JA, Parker HG, Ostrander EA. Breed-specific ancestry studies and genome-wide association analysis highlight an association between the MYH9 gene and heat tolerance in Alaskan sprint racing sled dogs. Mamm Genome. 2012:23(1-2):178–194. 10.1007/s00335-011-9374-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maier R, Flegontov P, Flegontova O, Işıldak U, Changmai P, Reich D. On the limits of fitting complex models of population history to f-statistics. eLife. 2023:12:e85492. 10.7554/eLife.85492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meadows JRS, Kidd JM, Wang GD. Genome sequencing of 2000 canids by the Dog10K consortium advances the understanding of demography, genome function and architecture. Genome Biol. 2023:24. 10.1186/s13059-023-03023-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Monteath AJ, Gaglioti BV, Edwards ME, Froese D. Late Pleistocene shrub expansion preceded megafauna turnover and extinctions in eastern Beringia. Proc Natl Acad Sci U S A. 2021:118(52):e2107977118. 10.1073/pnas.2107977118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moon KL, Huson HJ, Morrill K, Wang MS, Li X, Srikanth K, Zoonomia Consortium, Lindblad-Toh K, Svenson GJ, Karlsson EK, et al. Comparative genomics of Balto, a famous historic dog, captures lost diversity of 1920s sled dogs. Science. 2023:380(6643):eabn5887. 10.1126/science.abn5887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ní Leathlobhair M, Perri AR, Irving-Pease EK, Witt KE, Linderholm A, Haile J, Lebrasseur O, Ameen C, Blick J, Boyko AR, et al. The evolutionary history of dogs in the Americas. Science. 2018:361(6397):81–85. 10.1126/science.aao4776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pang JF, Kluetsch C, Zou XJ, Zhang AB, Luo LY, Angleby H, Ardalan A, Ekström C, Sköllermo A, Lundeberg J, et al. mtDNA data indicate a single origin for dogs south of Yangtze River, less than 16,300 years ago, from numerous wolves. Mol Biol Evol. 2009:26(12):2849–2864. 10.1093/molbev/msp195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patterson N, Moorjani P, Luo Y, Mallick S, Rohland N, Zhan Y, Genschoreck T, Webster T, Reich D. Ancient admixture in human history. Genetics. 2012:192(3):1065–1093. 10.1534/genetics.112.145037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perri AR, Feuerborn TR, Frantz LA, Larson G, Malhi RS, Meltzer DJ, Witt KE. Dog domestication and the dual dispersal of people and dogs into the Americas. Proc Natl Acad Sci U S A. 2021:118(6):e2010083118. 10.1073/pnas.2010083118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phillips CJ, Coppinger RP, Schimel DS. Hyperthermia in running sled dogs. J Appl Physiol. 1981:51(1):135–142. 10.1152/jappl.1981.51.1.135. [DOI] [PubMed] [Google Scholar]
- Pickrell J, Pritchard J. Inference of population splits and mixtures from genome-wide allele frequency data. PLoS Genet. 2012:8(11):e1002967. 10.1371/journal.pgen.1002967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pitulko VV, Kasparov AK. Archaeological dogs from the early Holocene Zhokhov site in the eastern Siberian Arctic. J Archaeol Sci Rep. 2017:13:491–515. 10.1016/j.jasrep.2017.04.003. [DOI] [Google Scholar]
- Plassais J, Parker HG, Carmagnini A, Dubos N, Papa I, Bevant K, Derrien T, Hennelly LM, Whitaker DT, Harris AC, et al. Natural and human-driven selection of a single non-coding body size variant in ancient and modern canids. Curr Biol. 2022:32(4):889–897.e9. 10.1016/j.cub.2021.12.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Potseluyeva Y.Historic and climatic prerequisites of appearance of the sled dog population of the Chukotka Peninsula shoreline. In: Beregovoy V, editor. The dogs of our ancestors. India: CATNUS Publishers; 2022. p. 914–918, 927–930. [Google Scholar]
- Ramos Madrigal J, Sinding MHS, Carøe C, Mak SS, Niemann J, Samaniengo Casruita JA, Fedorov S, Kandyba A, Germonpré M, Bocherens H, et al. Genomes of extinct Pleistocene Siberian wolves provide insights into the origin of present-day wolves. Curr Biol. 2021:31(1):1–12.e5. 10.1016/j.cub.2020.09.070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reich D, Thangaraj K, Patterson N, Price AL, Singh L. Reconstructing Indian population history. Nature. 2009:461(7263):489–494. 10.1038/nature08365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sams AJ, Boyko AR. Fine-scale resolution of runs of homozygosity reveal patterns of inbreeding and substantial overlap with recessive disease genotypes in domestic dogs. G3 Genes Genomes Genet. 2019:9(1):117–123. 10.1534/g3.118.200836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Savolainen P, Zhang YP, Luo J, Lundeberg J, Leitner T. Genetic evidence for an East Asian origin of domestic dogs. Science. 2002:298(5598):1610–1613. 10.1126/science.1073906. [DOI] [PubMed] [Google Scholar]
- Sinding MHS, Gopalakrishnan S, Ramos-Madrigal J, de Manuel M, Pitulko VV, Kuderna L, Feuerborn TR, Frantz LA, Vieira FG, Niemann J, et al. Arctic-adapted dogs emerged at the Pleistocene–Holocene transition. Science. 2020:368(6498):1495–1499. 10.1126/science.aaz8599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Skoglund P, Ersmark E, Palkopoulou E, Dalén L. Ancient wolf genome reveals an early divergence of domestic dog ancestors and admixture into high-latitude breeds. Curr Biol. 2015:25(11):1515–1519. 10.1016/j.cub.2015.04.019. [DOI] [PubMed] [Google Scholar]
- Stange M, Sánchez-Villagra MR, Salzburger W, Matschiner M. Bayesian divergence-time estimation with genome-wide SNP Data of Sea Catfishes (Ariidae) supports miocene closure of the Panamanian isthmus. Syst Biol. 2018:67(4):681–699. 10.1093/sysbio/syy006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sullivan PF, Meadows JR, Gazal S, Phan BN, Li X, Genereux DP, Dong MX, Bianchi M, Andrews G, Sakthikumar S, et al. Leveraging base-pair mammalian constraint to understand genetic variation and human disease. Science. 2023:380(6643):eabn2937. 10.1126/science.abn2937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomas B, Thomas P. Leonhard Seppala: the Siberian Dog and the golden age of Sleddog Racing 1908–1941. Missoula, Montana: Pictorial Histories Publishing Company; 2015. [Google Scholar]
- Thorsrud JA, Huson HJ. Description of breed ancestry and genetic health traits in Arctic sled dog breeds. Canine Med Genet. 2021:8(1):8–13. 10.1186/s40575-021-00108-z. [DOI] [PMC free article] [PubMed] [Google Scholar]