(вся статья переведена с использованием встроенного переводчика в Яндекс Браузере)
Аннотация
Изменчивость окружающей среды играет решающую роль в выборе места обитания для высших хищников, однако ее влияние в условиях антропогенного воздействия остается неясным. С августа 2021 года по март 2024 года мы провели 911 визуальных наблюдений за тигровыми акулами (Galeocerdo cuvier) в Фувамуле, Мальдивы. Мы использовали метод фотоидентификации для определения отдельных особей тигровых акул и агломеративную иерархическую кластеризацию для разделения акул на группы в зависимости от места обитания. Мы применили обобщенные аддитивные модели к кластерам мест обитания и группам по стадиям жизненного цикла, чтобы проверить, влияют ли температура поверхности моря, фаза муссона, взаимодействие лунных и приливно-отливных явлений, а также показатели ветра и волн на численность акул на дайв-сайте «Тигровая бухта». Модели объяснили 24–38 % отклонений и показали, что оседлые, относительно крупные самки предпочитают теплые температуры и муссонные циклы, в то время как мигрирующие, относительно мелкие акулы больше реагируют на погодные условия, направление ветра и взаимодействие лунных и приливно-отливных явлений. Наши результаты показывают, что естественные природные ритмы по-прежнему влияют на появление тигровых акул в местах их обитания даже при регулярном кормлении. Это согласуется с выводами, сделанными на основе наблюдений за другими скоплениями акул, которых подкармливают.
Введение
Численность хищных акул во всем мире сокращается в основном из-за чрезмерного вылова (Dulvy et al. 2021). Отсутствие таких хищников может привести к изменению структуры пищевой сети и функционирования экосистемы (Baum and Worm 2009). Однако присутствие акул обусловлено сложным сочетанием антропогенных, экологических и природных факторов (Heupel and Simpfendorfer 2014; Schlaff et al. 2014; Hammerschlag et al. 2022). Температура, глубина, приливно-отливные режимы, фазы Луны и сезонная изменчивость были определены как основные факторы, влияющие на перемещения акул и скатов (Schlaff et al. 2014). Температура оказывает сильное влияние на метаболизм акул и приводит к суточным, сезонным и долгосрочным изменениям в их ареале (Bernal et al. 2012; Sims et al. 2006; Ketchum et al. 2014; Osgood et al. 2021). Кроме того, поведенческая терморегуляция связана с размножением и добычей пищи: акулы ищут оптимальные температурные условия для максимальной эффективности метаболических процессов (Sims et al. 2006; Speed et al. 2012; Watanabe et al. 2021). Приливные циклы влияют на использование среды обитания такими видами, как тупорылые акулы Carcharhinus leucas, чернопёрые рифовые акулы C. melanopterus и лимонные акулы Negaprion brevirostris (Heupel et al. 2010; Lea et al. 2020), а в сочетании с фазами Луны — по крайней мере у девяти видов пластиножаберных (Andrzejaczek et al. 2024).
Тигровые акулы Galeocerdo cuvier — высшие хищники, играющие ключевую экологическую роль в самых разных морских средах обитания. Являясь ключевыми видами, они формируют популяции своей добычи и способствуют взаимодействию прибрежных и пелагических экосистем (Papastamatiou et al. 2015; Vaudo et al. 2014). Тигровые акулы были связаны с экологическими эффектами во многих морских местообитаниях, включая потенциальное влияние на динамику травоядных и структуру среды обитания, хотя эти эффекты зависят от контекста (Дедман и др., 2024). Эти акулы демонстрируют весьма изменчивые модели передвижения, охватывающие различные пространственные и временные масштабы, при этом на поведение влияют градиенты окружающей среды, физиологические потребности и доступность добычи, поддерживая контекстно-зависимую экологическую структуру (Lubitz et al. 2022).
В западной части Индийского океана (ЗЧИО) тигровые акулы демонстрируют как привязанность к определенным местам, так и способность к дальним миграциям (Эберт и др., 2021; Хейтхаус и др., 2007). В таких регионах, как La Réunion, Южная Африка и Мадагаскар, тигровые акулы демонстрируют сезонные изменения в пространственном поведении и сроках родов в зависимости от пола (Жакеман и др., 2013; Блейсон и др., 2015; Дикен и др., 2016). В Западной Австралии также наблюдаются сезонные колебания: чаще всего они встречаются в период с сентября по май, что, вероятно, связано с обилием добычи в местах кормежки, таких как Шарк-Бей (Heithaus 2001; Simpfendorfer et al. 2001; Wirsing et al. 2006). Эти закономерности показывают, как тигровые акулы реагируют на изменения окружающей среды и доступность ресурсов в Индийском океане (Ferreira et al. 2015); однако таких данных нет по Мальдивам — центральному, но малоизученному региону, несмотря на то, что он объявлен заповедником для акул (Vossgaetter et al. 2024).
В последние годы кормление акул стало распространенным и прибыльным занятием в туристических целях, приносящим значительный доход местной экономике (Галлахер и Хаммершлаг 2011; Сиснерос-Монтемайор и др. 2013). Однако это занятие может быть опасным как для людей, так и для акул (Брена и др. 2015; Галлахер и др. 2015). Снабжение акул пищевыми аттрактантами относится к использованию пищевых аттрактантов для увеличения вероятности встреч с акулами, хотя встречи остаются переменными и не гарантированы (Meyer et al., 2009). Места заготовки запасов обеспечивают контролируемую основу для проведения научных исследований и мониторинга внешних факторов, которые могут повлиять на вероятность встреч с морской фауной в небольшом пространственном масштабе. Измененные человеком экосистемы, в которых пища предсказуемо доступна круглый год, как, например, в Фувамуле на Мальдивах, потенциально могут изменить влияние природных факторов на распространение акул (например, китовых акул Rhincodon typus, Legaspi et al. 2020). В Фувамуле круглый год обитают половозрелые самки, а также молодь и иногда взрослые самцы (Sulikowski et al. 2024; Vossgaetter et al. 2024).
Оценка влияния факторов окружающей среды на тигровых акул, которые круглый год получают корм в Фувамуле, крайне важна для понимания того, как эти факторы влияют на их пространственную экологию, несмотря на постоянное поступление корма. Поскольку добыча акул для получения корма становится все более распространенным явлением во всем мире, исследования, предоставляющие данные о параметрах, влияющих на перемещение, распространение и поведение акул, независимо от промысла, крайне важны для управления целевыми видами. В настоящее время тигровых акул отлавливают по всему миру в нескольких местах (Séguigne et al. 2023), хотя условия окружающей среды, которые могут влиять на вероятность встречи с акулами в этих местах, часто остаются без внимания.
В местах, где акулы чувствуют себя комфортно, отдельные особи часто демонстрируют неоднородное поведение: одни возвращаются на одно и то же место в течение длительного времени, а другие появляются лишь время от времени. Такая вариативность в привязанности к месту может быть связана с различиями в стратегии поиска пищи, репродуктивном состоянии или терпимостью к местным условиям, а также влиять на то, насколько сильно факторы окружающей среды влияют на частоту встреч с акулами. Таким образом, разделение особей по характеру пребывания на одном месте может помочь определить, различаются ли реакции на окружающую среду у акул с разными стратегиями использования мест обитания.
Здесь мы ставим своей целью (а) исследовать влияние, если таковое имеется, переменных окружающей среды на численность тигровых акул в Фувамуле на Мальдивах; (б) и проверить, различаются ли экологические предикторы наблюдений за тигровыми акулами между группами особей с разной точностью определения местоположения, определенной на основе историй встреч с фотоидентификацией, и между стадиями жизни (взрослые и молодь).
Методы
Исследуемый объект
Фувамула — это атолл, состоящий из одного острова, расположенный в южной части Мальдивского архипелага, почти в 30 км к югу от экватора, в окружении Индийского океана (широта -0,30°, долгота 73,43°). Остров представляет собой океанический платформенный риф, окруженный окаймляющим рифом, и, в отличие от других атоллов Мальдив, не имеет лагуны. На юго-восточной оконечности острова находится 2-километровый отросток рифа (на местном языке — Фарикиде), представляющий собой умеренно наклонный коралловый риф (глубина 0–20 м) с обрывом. Батиметрия вблизи атолла обычно колеблется от 30 до 80 м глубины, хотя абиссальная равнина достигает глубин > 2000 м. Метеорологические данные свидетельствуют о наличии двух преобладающих муссонных фаз, северо-восточного муссона (декабрь-апрель) и юго-западного муссона (июнь-октябрь), которые чередуются с межсезонными муссонами в мае и ноябре (Шотт и др., 2009; Су и др., 2021), и смешанного полусуточного приливного режима (максимальная дальность 1,1 м) (UHSLC, Caldwell et al . 2015). Наш исследовательский участок расположен за пределами гавани, в месте для дайвинга под названием «Тигровая гавань». Это место для дайвинга (ранее описанное в работах Vossgaetter et al. 2024 и Reinero et al. 2025) находится на мелководье (< 11 м), где тигровые акулы добывают головы тунца или рыбные отбросы, спрятанные под камнями, ежедневно, круглый год, в среднем по 30-40 минут за погружение. Все съемки проводились в одном и том же месте заготовки, и водолазы оставались в постоянном диапазоне малых глубин (<11 м) в течение каждого периода наблюдения, обеспечивая стандартизированную глубину съемки в течение нескольких дней отбора проб (рис. 1).
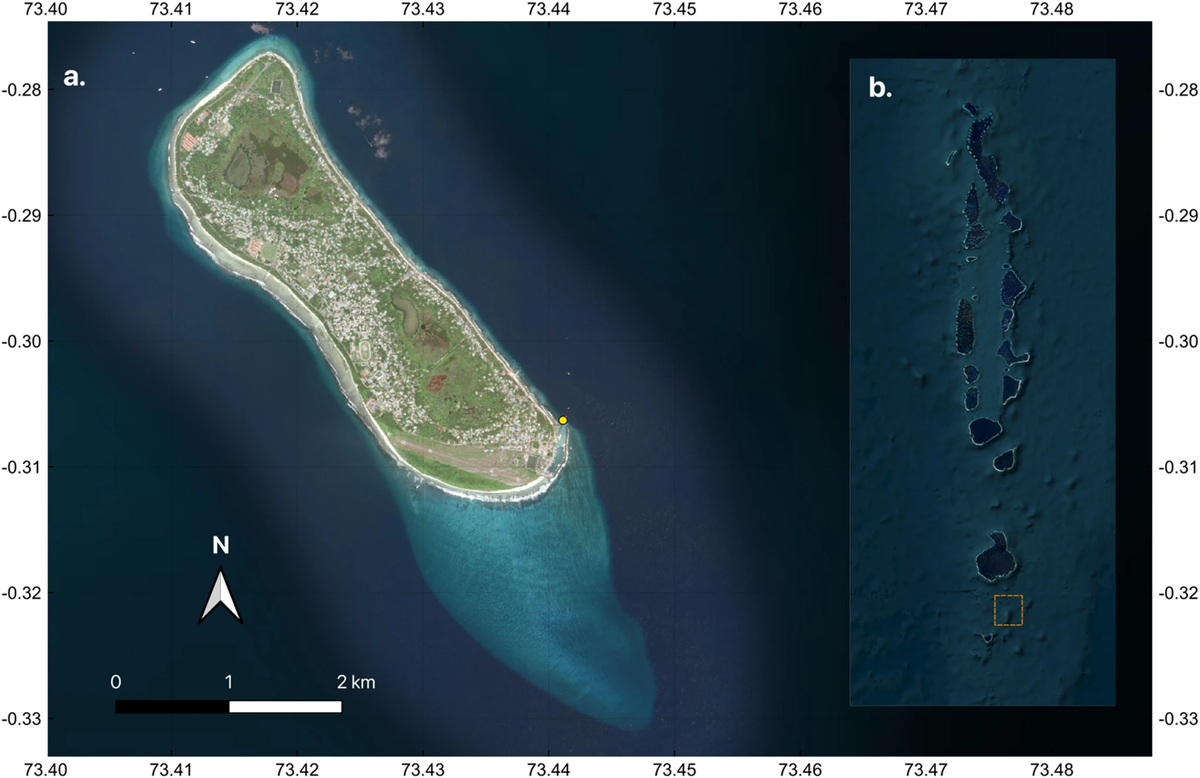
Сбор данных
Исследования проводились в период с августа 2021 по март 2024 года и включали в себя в общей сложности 877 погружений. Отбор проб проводился круглый год во все месяцы, с ежемесячным объемом погружений от 29 погружений (июнь) до 114 погружений (ноябрь), охватывающих четыре календарных года (2021-2024). Наблюдения за тигровыми акулами систематически регистрировались местными морскими биологами в соответствии с методологией, принятой Воссгеттером и др. (2024). Уникальные особи были идентифицированы на основе отличительных физических характеристик, которые сохранялись на протяжении всего исследования и позволяли избежать двойного учета (Vossgaetter et al. 2024).
В качестве переменной отклика использовалось количество уникальных тигровых акул, идентифицированных по фотографии, за календарный день во время стандартизированных погружений с целью сбора провизии. Погружения проводились ежедневно в одном и том же месте по единому протоколу, включавшему стандартную продолжительность (55–60 минут) и использование одной и той же приманки — голов тунца. Подсчет акул велся с момента подачи приманки до окончания погружения. Поскольку место, продолжительность и способ подачи приманки были одинаковыми во все дни сбора данных, ежедневные подсчеты акул можно было считать сопоставимыми без дополнительной корректировки.
Жизненный цикл (молодые особи или взрослые) определялся в соответствии с размерными категориями, которые использовали Воссгеттер и соавторы (2024) при описании тигровых акул в той же популяции у атолла Фувамула, на основе оценки общей длины по фотоидентификации и фотограмметрии. Условия окружающей среды отслеживались с помощью комбинации данных, полученных непосредственно на месте, и данных дистанционного зондирования (см. таблицу 1).
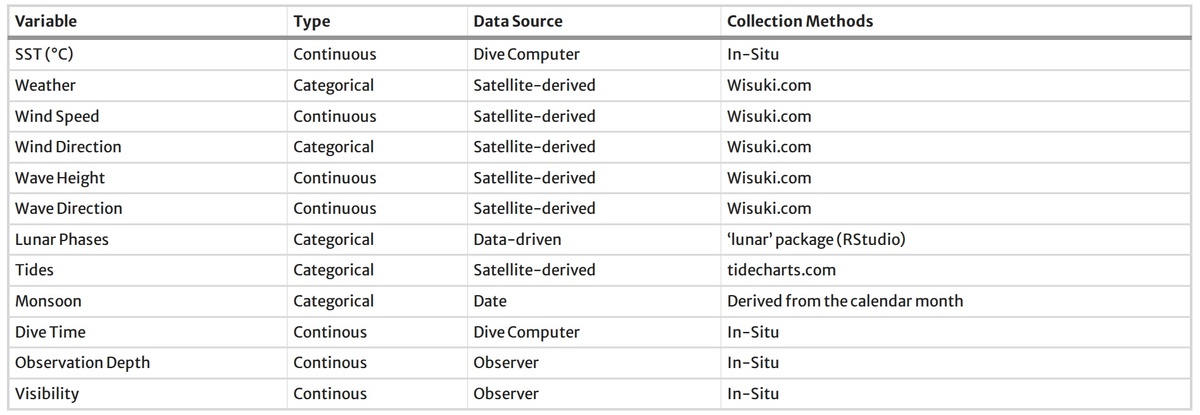
Температура поверхности моря (ТПМ, °C) измерялась с помощью откалиброванных подводных компьютеров, а скорость и направление ветра, а также высота и направление волн — с помощью Wisuki.com, в соответствии с тем же подходом, что и в работе Ionuzzi et al. (2023). Данные о погоде собирались на месте с использованием категориальной классификации погодных условий с четырьмя уровнями: (а) ‘Солнечно’, ‘Дождь’, ‘Облачно’, ‘Переменная облачность’’.
Фазы Луны были определены с помощью пакета `lunar` в R (версия 2024.12.1 + 563) и сопоставлены с соответствующей датой, что позволило синхронизировать данные об окружающей среде с лунным циклом, который может влиять на поведение акул (RStudio, 2024).
Кластеры, отражающие привязанность к месту обитания
Чтобы оценить влияние факторов окружающей среды на группы акул, мы сгруппировали особей с разной степенью привязанности к месту обитания, чтобы выявить возможные закономерности на индивидуальном уровне. Анализ проводился на основе глобального индекса привязанности к месту обитания (Site Fidelity Index, SFI), который использовали Сегень и соавторы (2023). SFI рассчитывался как доля погружений за период наблюдения, во время которых была замечена акула, и определялся путем деления количества погружений, во время которых была замечена акула, на общее количество погружений. Значения варьируются от 0 до 1, где более низкий показатель SFI указывает на меньшую привязанность к исследуемой территории, а более высокий — на большую.
На основе индивидуальных значений SFI был проведен агломеративный иерархический кластерный анализ (AHC) для разделения акул со схожими показателями оседлости. Несходство между особями рассчитывалось с помощью евклидова расстояния, а в качестве алгоритма кластеризации использовался метод Уорда. Кластеризацию проводили с помощью функции hclust из пакета R stats с параметром method = “ward.D2” (Murtagh и Legendre 2014) по методике Кауфмана и Руссева (1990) и Legendre и Legendre (2012). Оптимальное количество кластеров определяли с помощью метода среднего силуэта, реализованного в пакете R cluster (Maechler et al. 2021).
Обобщенные аддитивные модели (ОАМ)
Чтобы выяснить, как факторы окружающей среды и временные переменные влияют на ежедневную встречаемость тигровых акул, мы смоделировали количество уникальных тигровых акул, замеченных за календарный день, в зависимости от факторов окружающей среды. Каждое наблюдение представляло собой одно стандартизированное погружение с целью сбора образцов, проведенное в одном и том же месте, на одной и той же глубине и в течение одного и того же времени, что обеспечивало сопоставимость результатов погружений. Поэтому в модели не включался параметр смещения. Учитывая количественный характер зависимой переменной (количество отдельных тигровых акул за календарный день) и экологических предикторов, для анализа данных были использованы обобщенные аддитивные модели (ОАМ) с помощью пакета mgcv (Вуд 2017) в RStudio. Для оценки влияния экологических и временных факторов на численность акул использовалось распределение Пуассона и логарифмическая связь. Все предикторы были включены в модель в качестве параметрических членов (то есть не использовались сплайновые сглаживатели), поэтому размерность базиса (узлов) не применялась. Соответственно, не применялась и диагностика, специфичная для сглаживающих членов (например, проверка размерности базиса и конгруэнтности). Соответствие модели оценивалось с помощью диагностики остатков и проверки на сверхдисперсию, а коллинеарность предикторов — с помощью коэффициентов увеличения дисперсии. В качестве исследовательских переменных были включены экологические и временные факторы (например, высота волн в метрах, направление ветра по сторонам света, температура воды в градусах Цельсия, восемь уникальных лунных фаз, категория приливов, сезон муссонов и погодные условия; см. таблицу 1). Выбор модели начался с полной модели, включающей все возможные предикторы. Предикторы последовательно исключались методом обратного пошагового отбора, при этом на каждом этапе модели сравнивались с помощью информационного критерия Акаике (AIC) (Papastamatiou et al. 2015). Окончательная модель сохранила комбинацию предикторов , которые минимизировали AIC . После выбора наиболее подходящей модели мы ввели отдельную модель, включающую термин взаимодействия прилива и фазы Луны, чтобы проверить, различается ли влияние прилива на количество тигровых акул в разные фазы Луны. Коллинеарность между предикторами оценивалась с использованием коэффициентов инфляции дисперсии, а соответствие модели оценивалось с помощью остаточной диагностики (т. Е. Остатков по сравнению с подобранными значениями, коэффициентов избыточного распределения), чтобы убедиться, что допущения были выполнены (Ackerman et al., 2000). Были построены отдельные модели для (i) кластеров местообитаний, выявленных с помощью иерархической кластеризации, и (ii) категорий жизненных стадий (взрослые особи и молодняк), чтобы оценить, различаются ли факторы окружающей среды в зависимости от привязанности к месту обитания и онтогенетических классов.
Результаты
В ходе агломеративной иерархической кластеризации (АХК) были выделены два отдельных кластера: в кластер 1 вошли 30 человек, а в кластер 2 — 190 человек. Пороговым значением АХК была установлена высота 1,05, а коэффициент когенетической корреляции составил 0,845, что указывает на хорошее представление лежащих в основе различий.
Кластеризация была основана на значениях индекса точности сайта (SFI), безразмерного индекса в диапазоне от 0 до 1, с более высокими значениями, указывающими на более высокую точность сайта. Кластер 1 демонстрировал значительно более высокие значения SSFI (среднее значение ± SD = 0,1698 ± 0,0428) и состоял полностью из женщин (100% женского пола; 73,3% классифицированы как взрослые). Многие особи в этой группе были замечены беременными (Суликовски и др., 2024). В дальнейшем этот кластер будет называться «высоко резидентным» (HR).
Во втором кластере значения SFI были значительно ниже (среднее ± стандартное отклонение = 0,0269 ± 0,0307), в том числе у особей, замеченных всего один раз за весь период исследования. В этот кластер входили как молодые, так и взрослые особи, преимущественно самки (82,1 % самок; 51,6 % взрослых особей). В дальнейшем эту группу будем называть «менее оседлой» (МО).
Обобщённые аддитивные модели (ОАМ) объяснили от 24,3% до 38,2% отклонений в данных о наблюдениях за тигровыми акулами, при этом скорректированные значения r² варьировались от 0,188 до 0,232 (таблица 2). Значимые предикторы различались в зависимости от места обитания и жизненного цикла.
Для тигровых акул (кластер 1, таблица 2) модель объяснила 29,6 % отклонений (r² = 0,208). Количество наблюдений увеличивалось с ростом температуры поверхности моря (p = 0,019). Ветер с востока-северо-востока (ENE), севера (N) и юго-востока (SE) также оказывал значительное влияние на количество наблюдений (p = 0,014, p = 0,016,P = 0,008). Наблюдались эффекты, связанные с фазами Луны и приливами и отливами: количество наблюдений увеличивалось в полнолуние и во время прилива (p = 0,04).
Для тигровых акул (кластер 2 — таблица 2) модель объяснила 38,2 % отклонений (r² = 0,232). Наблюдения положительно коррелировали с северо-восточным муссоном (p = 2,9e-07), дождливой (p = 0,018) и солнечной (p = 0,04) погодой, а также с поверхностными ветрами с юга (S) (p = 0,008), юго-запада (SW) (p = 0,017), запада (W) (p = 0,006) и западо-юго-запада (WSW) (p = 0,0019). Значительное лунно-приливное взаимодействие было обнаружено во время прилива и убывающей фазы Луны (p = 0,0118, p = 0,023).
Для молоди акул (таблица 2) модель объяснила 24,3 % отклонений (r² = 0,188). Количество наблюдений положительно коррелировало с полнолунием (p = 0,03), фазой муссона (p = 9,2e-06) и средней температурой поверхности моря (p = 0,009). Повышенная видимость также наблюдалась в частично облачную (p = 0,017) и дождливую (p = 0,047) погоду, а также при ветре с запада-юго-запада (WSW) (p = 0,031). Влияние Луны и приливов было значительным во время прилива (p = 0,02) и в первую четверть лунного цикла.
Для взрослых акул (таблица 2) модель объяснила 25 % отклонений (r² = 0,197). На количество наблюдений влияли поверхностная температура воды (p = 1,2e-05) и полнолуние (p = 0,008). Сезон муссонов (p = 0,001) и направление ветра с востока-юго-востока (ESE) (p = 0,02), северо-востока (NE) (p = 0,03) и юго-юго-востока (SSE) (p = 0,04) также были значимыми прогностическими факторами. Взаимодействие лунных и приливно-отливных явлений наблюдалось во время прилива и убывающей луны (p = 0,043).
Влияние сезонных и температурных условий на наблюдения за тигровыми акулами
Температура поверхности моря (ТПМ) оказалась стабильным положительным фактором, влияющим на количество наблюдений за тигровыми акулами во всех четырех группах (рис. 2; табл. 2). SST был значимо и положительно связан с наблюдениями за акулами у особей с высоким уровнем резидентности (Оценка = 0,0739, SE = 0,0316, z = 2,3350, p = 0,0195), менее резидентных особей (оценка = 0,0507, SE = 0,0208, z = 2,4347, p = 0,0149), молоди (оценка = 0,0799, SE = 0,0308, z = 2,5952, p = 0,0095) и взрослых особей (оценка = 0,0917, SE = 0,0210, z = 4,3763, р < 0,001). В совокупности эти закономерности указывают на увеличение количества наблюдений при более высоких температурах поверхности независимо от места обитания и стадии развития.
Аналогичным образом, муссонные фазы также стали факторами воздействия на наблюдения за тигровыми акулами, причем количество наблюдений во время северо-восточного муссона во всех группах было неизменно выше (рис. 3). Эффект был наиболее сильным среди взрослых акул (оценка = -0,55, SE = 0,12, z-балл = -4,60, p < 0,001) и особей с высоким уровнем оседлости (оценка = -0,47, SE = 0,13, z-балл = -3,56, p < 0,001), что указывает на значительно меньшее количество наблюдений во время юго-восточного муссона по сравнению с северо-восточным. У менее оседлых акул наблюдалась значительная сезонная реакция (оценка = -0,42, стандартная ошибка = 0,14, z = -2,97, p = 0,003), в то время как у молоди реакция была слабее и статистически незначима (оценка = -0,26, стандартная ошибка = 0,17, z = -1,53, p = 0,126) (таблица 2). В целом эти результаты свидетельствуют о сезонном распределении тигровых акул в районе исследования, при этом в период северо-восточного муссона их особенно много среди оседлых особей и взрослых акул.
Обсуждение
Общие тенденции
Это исследование выявило чёткие временные закономерности в наблюдениях за тигровыми акулами в Фувамуле. Наши результаты показывают, что сочетание сезонных и краткосрочных факторов окружающей среды объясняет значительную часть вариативности частоты наблюдений. Акулы в бухте Тайгер-Харбор не появляются там случайно, а реагируют на внешние факторы, даже в этом месте нагула. Фазы муссонов и температура поверхности моря оказались ключевыми факторами, влияющими на все стадии жизни и группы оседлости акул. Тенденции, выявленные с помощью GAM, свидетельствуют о том, что тигровые акулы корректируют частоту посещений в зависимости от сезонных изменений и более мелких циклов, что, вероятно, является частью их поведенческой стратегии, направленной на оптимизацию питания и его соответствие физиологическим потребностям, а не просто следствием наличия кормовой базы. Как и в случае с другими видами, популяции акул могут динамически реагировать на окружающую среду, выбирая места обитания в зависимости от контекста (Papastamatiou et al. 2015; Lubitz et al. 2022).Наши результаты свидетельствуют о более выраженном сезонном характере распространения, обусловленном фазами муссонов, температурой поверхностного слоя воды и лунно-приливными циклами, которые существенно влияют на распределение акул, при этом наблюдаются заметные различия между стадиями жизненного цикла и группами оседлости. Различия между группами могут также отражать стратегии перемещения, характерные для разных стадий жизненного цикла: более непостоянные молодые особи реагируют на более масштабные изменения окружающей среды, в то время как взрослые особи, ведущие оседлый образ жизни, сильнее реагируют на локальные изменения.
Сезонность и муссоны
Ярко выраженный сезонный сигнал был очевиден при наблюдениях тигровых акул, тесно связанных с региональным циклом муссонов. Эта закономерность была подтверждена lдолгосрочные данные фотографической идентификации с того же участка, где самцы почти полностью отсутствовали с июля по сентябрь, а пик наблюдений за самками пришелся на ноябрь-апрель (Vossgaetter et al., 2024). Сезонное снижение количества наблюдений во время юго-западного муссона свидетельствует о том, что часть популяции, особенно самцы и, возможно, самки на последних сроках беременности, мигрируют или реже посещают это место. По данным Бетцлера и др. (2018), северо-восточный муссон приносит на Мальдивы более спокойные и часто более прозрачные воды с другими течениями. Такие условия могут максимально расширить возможности для кормежки и способствовать более активному скоплению акул в одном месте. Такэмото и соавторы (2014) обнаружили, что муссоны влияют на рифовые и пелагические пищевые сети, благодаря чему в определенные месяцы может увеличиваться количество рыб, служащих добычей, или появляться больше возможностей для падальщиков. Тигровые акулы — оппортунистические хищники (Лоу и соавторы. 1996), и они могут выбирать время для пребывания в Фувахмуле так, чтобы оно совпадало с периодами, когда добыча пищи наиболее выгодна, а также с периодами, когда океан более спокоен, что позволяет им эффективно патрулировать территорию и взаимодействовать с дайверами.
С другой стороны, юго-западный муссон характеризуется более сильными ветрами, обильными осадками и в целом более бурными морскими условиями, а также снижением доступности пищи из-за низкого туристического сезона. Из-за этих условий акулы могут искать альтернативные места или более защищённые от непогоды места обитания, что приводит к уменьшению количества наблюдений в местах нагула. На Багамах существует гипотеза, что тигровые акулы могут использовать места нагула таким образом, что нагульные мероприятия сами по себе не являются единственной причиной, по которой они ограничивают использование среды обитания и перемещаются только к местам нагула. Другие факторы могут объяснить особенности расселения и использования пространства в местах с высокой посещаемостью, подверженных влиянию акульего туризма, такие как предпочтения в отношении добычи и ее доступность, а также размножение (то есть беременность и роды) (Хаммершлаг и др. 2012, 2017). Аналогичные закономерности наблюдались и в местах кормления акул на Фиджи (Брунсвайлер и Барнетт 2013) и во Французской Полинезии (Сегень и др. 2023), где изменчивость окружающей среды по-прежнему влияет на присутствие акул, несмотря на регулярное кормление.
Наблюдаемая сезонная закономерность в местах обнаружения тигровых акул согласуется с сезонными изменениями среды обитания, зафиксированными у других крупных пластиножаберных рыб этого региона. Например, китовые акулы на Мальдивах перемещаются в зависимости от муссонов, переходя с одной стороны атоллов на другую по мере смены направления муссонных течений, и собираются там, где в каждый сезон наблюдается цветение планктона (Anderson and Ahmed 1993). На атолле Южный Ари китовые акулы и скаты манта перемещаются между восточной и западной рифовыми зонами в зависимости от динамики сезонного планктона, эффективно отслеживая колебания продуктивности, обусловленные муссонным циклом (Anderson et al. 2011; Valsecchi et al. 2021).
Еще одним фактором является сезонная привязанность крупных акул к определенным местам, связанная с репродуктивными циклами (Hammerschlag et al. 2011). Многие виды акул совершают сезонные миграции или меняют среду обитания в зависимости от времени спаривания или рождения детенышей (Heupel et al. 2015). Например, в западной части Атлантического океана взрослые самки тигровых акул мигрируют в зависимости от сезона, предположительно для того, чтобы родить или спариться в определенных районах в определенное время (Hammerschlag et al. 2012; Sulikowski and Hammerschlag 2023). В нашем исследовании сокращение количества наблюдений (в середине года) приходится на конец весны и лето. Возможно, в этот период беременные самки покидают места скопления, чтобы родить, если существует синхронный сезон размножения (Кастро 2009). Таким образом, снижение численности в сезон муссонов можно объяснить тем, что самки, готовящиеся к родам, уходят в отдалённые места для размножения, а некоторые возвращаются с северо-восточным муссоном в конце года. Этот вывод подтверждается недавним исследованием, проведенным в Фувахмуле (Суликовский и др., 2024), в ходе которого в мае, за несколько месяцев до начала юго-западного муссона, было обнаружено множество беременных самок на разных стадиях беременности. Хотя для подтверждения таких перемещений потребуются данные прямого отслеживания, наблюдаемая нами ярко выраженная сезонность хорошо согласуется с гипотезой о том, что тигровые акулы циклически используют это место в зависимости от изменений окружающей среды, вызванных муссонами, и особенностей их жизненного цикла.
Температура
Среди изученных факторов окружающей среды температура поверхности моря оказалась наиболее значимым фактором, влияющим на появление тигровых акул, особенно среди акул, ведущих оседлый образ жизни. При повышении температуры воды количество наблюдений заметно увеличивалось, что позволяет предположить, что эти пойкилотермные акулы выбирают температурные условия, соответствующие их жизненным потребностям.
Известно, что температура является ключевым фактором, определяющим передвижение акул и выбор среды обитания, поскольку она напрямую влияет на скорость метаболизма и энергетический баланс (Bernal et al. 2012; Schlaff et al. 2014). Многочисленные исследования показали, что акулы часто демонстрируют поведенческую терморегуляцию, перемещаясь в зоны с предпочтительной температурой, чтобы оптимизировать биологические функции и процессы, связанные с жизнедеятельностью (Sims et al. 2006; Araujo et al. 2020; Pillans et al. 2021). В нашем исследовании положительная корреляция между температурой и частотой обнаружения акул, вероятно, объясняется тем, что в более теплые периоды акулы собираются в прибрежных водах острова, что связано с возможностью воспользоваться метаболическими преимуществами более высокой температуры воды. Кроме того, в более теплой воде ускоряются такие процессы, как пищеварение и, что особенно важно, беременность у акул, за счет повышения скорости метаболизма и эмбрионального роста (Speed et al. 2012; Watanabe et al. 2021). Мы действительно заметили, что в группе акул с наибольшей привязанностью к месту обитания преобладали половозрелые самки, многие из которых, по данным Суликовски и др. (2024), были беременны. Недавние неинвазивные ультразвуковые исследования в этом месте обнаружили эмбрионы у 93% всех обследованных половозрелых самок, что является прямым доказательством того, что это место служит местом вынашивания тигровых акул (Sulikowski et al., 2024). Учитывая, что у живородящих тигровых акул длительный период беременности (~12–16 месяцев) и относительно низкая репродуктивная способность, способность к терморегуляции за счет выбора мест с теплой водой может служить адаптивной стратегией для повышения репродуктивного успеха, как это наблюдается у других видов акул (Хайт и Лоу 2007; Матич и др. 2015), или для восстановления температуры тела после погружения в более прохладные воды на глубине.
Интересно, что влияние температуры было наиболее выражено у взрослых акул и в меньшей степени у молоди, что указывает на значительные различия в использовании пространства, связанные с тепловыми предпочтениями. У более крупных самок может быть больший стимул выбирать более теплые места обитания из-за требований беременности, в то время как молодь может быть более терпимой к диапазону температур или может более широко перемещаться в поисках пищи (Sulikowski et al., 2016). В Фувамуле взрослые самки могли использовать это место как своего рода «ясли» для развития потомства, оставаясь в комфортном температурном диапазоне. Наличие двух отдельных эмбриональных групп, обнаруженных с помощью ультразвука (Суликовски и др., 2024), позволяет предположить, что разные самки могут находиться на разных стадиях беременности, что, возможно, указывает на асинхронный или поэтапный репродуктивный цикл в популяции. Некоторые самки могут находиться в середине беременности, в то время как другие приближаются к родам, что может объяснить, почему не все взрослые самки покидают это место одновременно. Такая гибкость согласуется с появляющимися данными о том, что репродуктивные циклы тигровых акул могут различаться в зависимости от региона и особи (Уитни и др. 2007; Мануцци и др. 2022), а не все акулы, живущие по строго синхронизированному двухгодичному циклу. Вполне вероятно, что Фувахмулах — это круглогодичное безопасное убежище с оптимальной температурой, где беременные самки, живущие по разным циклам, могут оставаться до тех пор, пока им не нужно будет уплыть, чтобы родить. После родов самки могут вернуться на это место, чтобы восстановить силы в теплых, богатых пищей водах или снова спариться и вынашивать потомство, если их репродуктивный цикл короче.
Интерпретация взаимосвязи между температурой и встречами с акулами имеет ряд ограничений. Во-первых, наши данные о температуре отражают условия на поверхности во время погружений. У нас не было возможности непрерывно регистрировать температуру на месте и получать информацию о температуре на глубине. Возможно, выбор микросреды обитания мог повлиять на температуру, которую ощущали акулы, но мы не смогли это выяснить. Однако, учитывая, что большинство случаев взаимодействия акул с дайверами происходило на относительно небольшой глубине, измеренная температура, скорее всего, отражает условия, влияющие на поведение акул в этом месте. Во-вторых, хотя заманчиво сделать вывод, что более теплая вода вызывает большее присутствие акул, возможно также, что частично в этом виноват какой-то другой фактор, коррелирующий с сезонной температурой, такой как связанные с муссонами изменения течений или добычи. Мы попытались учесть сезон муссонов в моделях, и температура по-прежнему оставалась сильным независимым предиктором, подтверждающим истинные тепловые предпочтения. Кроме того, социальные сигналы и иерархия на индивидуальном уровне могут быть ответственны за меньшее присутствие несовершеннолетних на участке круглый год (Воссгеттер и др., 2024). У более крупных самок тигровых акул уровень тестостерона выше, и они доминируют в местах кормежки на Багамских островах (Rangel et al. 2022). Аналогичным образом на Фувахмуле более крупные взрослые самки оказывают влияние на более мелких сородичей и доминируют в местах кормежки (Reinero et al. 2025), что приводит к уменьшению количества наблюдений за молодью в присутствии взрослых особей и объясняет менее выраженное влияние факторов окружающей среды на молодь.
Тем не менее для подтверждения активного терморегуляторного поведения было бы полезно провести экспериментальные исследования или наблюдения. Наконец, мы признаем, что кормление может искажать естественные закономерности: если акул кормить, они могут переносить неоптимальные температуры. Тот факт, что они все равно предпочитали более теплые условия, указывает на то, что даже при кормлении их врожденная терморегуляция является движущей силой. Это открытие подчеркивает важность учета физиологических факторов, таких как температура, при организации мест скопления акул, особенно тех, которые являются критически важными для размножения.
Влияние приливов и отливов
Наш анализ выявил слабое, но статистически значимое влияние фазы прилива на количество наблюдений за акулами. В целом тигровых акул чаще замечали во время определенных приливов, особенно во время прилива или в полнолуние, о чем свидетельствуют несколько значимых взаимодействий между приливами и фазами Луны (например, увеличение количества наблюдений во время прилива в полнолуние и в полнолуние на убывающей Луне). Небольшое влияние приливов и отливов неудивительно, учитывая особенности нашего исследовательского участка. Тигровая бухта — это мелководный рифовый склон вблизи экватора, где амплитуда приливов и отливов невелика (порядка 1 м или меньше), а геоморфология не способствует возникновению сильных приливных течений, в отличие от устьев рек и лагун, где приливы могут резко менять глубину и заставлять акул перемещаться, как показали исследования Акермана и др. (2000). В совокупности эти условия могут позволить акулам оставаться в этом районе при любом уровне прилива и отлива. Их предпочтения едва уловимы и, возможно, связаны с удобством плавания или перемещения добычи при слабом течении.
Предыдущие исследования показали, что приливно-отливные циклы могут влиять на поведение и среду обитания прибрежных акул (Andrzejaczek et al. 2024), особенно молоди, обитающей на мелководье. Например, молодые тупорылые акулы в устье реки перемещаются вместе с приливом, чтобы использовать затопленные участки, а рифовые акулы, такие как чернопёрые рифовые акулы и лимонные акулы, часто проявляют повышенную активность или встречаются чаще во время приливов, когда приплывает добыча (Wetherbee et al. 2007; Heupel et al. 2010; Lea et al. 2020). Несмотря на незначительную роль приливных циклов, стоит отметить взаимодействие приливов и фаз Луны. Сочетание определенной фазы луны и прилива (например, прилив во время первой четверти луны) привело к небольшому увеличению количества наблюдений за детенышами акул, что позволяет предположить, что время приливов относительно уровня освещенности в ночное время может влиять на активность акул. Многие рифовые организмы синхронизируют свою активность с лунным освещением и приливами, что потенциально создает для акул благоприятные условия для охоты, как показали исследования Хаммершлага и др. (2017). Однако, учитывая сложность выделения таких эффектов и необходимость проведения множества сравнений, мы призываем не придавать чрезмерное значение этим взаимодействиям.
Мы также признаем, что наш анализ приливов и отливов имеет некоторые ограничения, связанные с региональным масштабом прогнозов, из-за чего мы можем не учитывать мелкомасштабные изменения или сдвиги во времени в конкретном месте. Кроме того, погружения обычно проводились в светлое время суток из соображений безопасности. Таким образом, любые реакции акул на приливы и отливы в ночное время, когда наблюдателей не было на месте, не попали бы в наш набор данных. Например, если бы акулы подплывали ближе к берегу во время прилива, чтобы полакомиться падалью, мы бы не заметили эту закономерность. Несмотря на эти оговорки, включение приливных факторов в наиболее подходящие модели указывает на то, что даже в системах с низкой амплитудой тигровые акулы незначительно меняют своё поведение в зависимости от прилива. Таким образом, при организации природоохранных мероприятий или туристических операций не обязательно строго ориентироваться на таблицы приливов и отливов, хотя для безопасности дайверов разумно избегать сильных течений.
Гидродинамические условия
Несколько переменных, связанных с ветром и волнами, существенно влияли на появление тигровых акул, особенно в местах, где они не живут постоянно (кластер 2). Акул было больше, когда дул ветер с юга, юго-запада, запада и западо-юго-западного направления, что характерно для юго-западного муссона и приводит к возникновению течений на рифах (Lea et al. 2020). Такие ветры могут усиливать запах наживки или падали, привлекая акул в гавань. Напротив, акулы, ведущие оседлый образ жизни (кластер 1), реже попадались на глаза при восточных ветрах (северо-восточных, северо-восточных и юго-восточных), возможно, потому, что прибрежные течения могут распространять шлейфы запаха вдоль края рифа, делая их более заметными для патрулирующих акул и увеличивая частоту посещения мест, как предположили Папастаматиу и соавторы (2015).
Аналогичным образом высота и направление волн были незначительными, но в некоторых моделях оказывали существенное влияние, что позволяет предположить, что волнение на поверхности влияет на видимость и перемещение добычи. Учитывая, что в Тигровой бухте волноприбойная полоса маломощная, влияние волн было незначительным, но тем не менее оно способствовало мелкомасштабным колебаниям в численности акул.
Заключение
Настоящее исследование позволяет по-новому взглянуть на факторы окружающей среды, влияющие на встречи с тигровыми акулами в Фувамуле, Мальдивы. Несмотря на антропогенное воздействие, связанное с деятельностью по обеспечению жизнедеятельности на территории, на появление тигровых акул, очевидно, влияли естественные природные ритмы. Наши результаты свидетельствуют о том, что сезонные встречи с акулами в большей степени зависят от муссонов, температуры поверхностного слоя воды и лунно-приливных циклов, которые существенно влияют на их характер, при этом наблюдаются заметные онтогенетические различия между группами и стадиями жизни. На взрослых самок, предпочитающих обитать в одном месте, сильно влияла температура воды, что согласуется с их терморегуляторным поведением во время беременности. В то же время менее оседлые особи, в том числе молодь и взрослые особи, появляющиеся в одном и том же месте нерегулярно, в большей степени реагировали на смену муссонов, направление ветра и более общие погодные условия, что указывает на сезонное использование места обитания, связанное с поиском пищи или расселением. Такая разная реакция свидетельствует о том, что Фувахмулах предлагает идеальные условия для длительного пребывания самок, готовых к размножению, и временных гостей из других демографических групп, удовлетворяя многочисленные потребности жизненного цикла в рамках одной рифовой системы.
Благодарности
Авторы выражают признательность компании Pelagic Divers Fuvahmulah (PDF) и Fuvahmulah Dive School (FDS) за их поддержку в проведении исследований. В частности, мы благодарим Mr Ahmed Inah, Ms Tatiana Ivanova и Mr Ibrahim Zihad за их финансовый вклад в проведение этого исследования. Мы также благодарим рецензентов за замечания, которые помогли улучшить рукопись.
Финансирование
Финансирование открытого доступа стало возможным благодаря проекту Projekt DEAL. Авторы заявили, что финансовые источники для проведения полевых исследований были полностью независимыми, а в процессе подготовки рукописи не получалась никакая финансовая, грантовая или иная поддержка.
Информация об авторе
Натан Перишич и Филиппо Бокки в равной степени внесли свой вклад в эту работу.
Авторы и аффилированные лица
Fuvahmulah Dive School, Fuvahmulah Shark Research, Fuvahmulah, 18011, Republic of MaldivesNathan Perisic, Tatiana Ivanova & Abdullah Niyaz
- Pelagic Divers Fuvahmulah, Nature Friends of Maldives, NGO, Fuvahmulah, 18011, Republic of MaldivesFilippo Bocchi, Ahmed Inah & Ahmed Hassan
- Leibniz Centre for Tropical Marine Research, 28334, Bremen, GermanyLennart Vossgaetter
- Ministry of Tourism and Environment, Male’, Maafannu, 20392, Republic of MaldivesAhmed Shan
- Marine Research and Conservation Foundation, Lydeard St Lawrence, Somerset, UKGonzalo Araujo
- Environmental Science Program, Department of Biological and Environmental Sciences, College of Arts and Sciences, Qatar University, Doha, QatarGonzalo Araujo
Вклад
Натан Перишич — концептуализация, формальный анализ, исследование, методология, программное обеспечение, визуализация, написание — первоначальный черновик, рецензирование, редактирование.
Филиппо Бокки — концептуализация, формальный анализ, исследование, методология, программное обеспечение, визуализация, написание — первоначальный черновик, рецензирование, редактирование.
Леннарт Фоссгеттер — концептуализация, формальный анализ, исследование, методология, программное обеспечение, написание — рецензирование и редактирование.
Татьяна Иванова — поиск финансирования.
Абдулла Нияз — обработка данных.
Ахмед Инах — привлечение финансирования.
Ахмед Хассан — обработка данных,
Ахмед Шан — написание, рецензирование и редактирование.
Гонсало Араухо — концептуализация, методология, супервизия, визуализация, написание, рецензирование, редактирование.
Автор, ответственный за переписку
Переписка с Леннартом Фоссгеттером.
Этические декларации
Конфликт интересов
Авторы заявляют об отсутствии конфликта интересов, связанного с данной работой.
Этическая экспертиза
Этическая экспертиза не требовалась, поскольку это исследование проводилось в рамках обсервационного проекта. Исследование проводилось в соответствии с рекомендациями и на основании разрешений на проведение исследований, выданных Агентством по охране окружающей среды (ежегодно продлеваемое разрешение: EPA/2021/PA-F01) и Министерством рыболовства, морских ресурсов и сельского хозяйства Мальдивских островов (ежегодно продлеваемое разрешение: 30-D/PRIV/2021/190). Использованные методы были неинвазивными и не причинили вреда подопытным животным.
Дополнительная информация
Ответственный редактор: Э. Шнайдер.
Примечание издателя
Springer Nature сохраняет нейтралитет в отношении юрисдикционных претензий на опубликованные карты и институциональную принадлежность.
Права и разрешения
Открытый доступ Эта статья лицензирована по международной лицензии Creative Commons Attribution 4.0, которая разрешает использование, совместное использование, адаптацию, распространение и воспроизведение на любом носителе или в любом формате при условии, что вы отдаете должное автору (ам) и источнику, предоставляете ссылку на лицензию Creative Commons и указываете, вносились ли изменения. Изображения или другие материалы третьих лиц, содержащиеся в этой статье, включены в лицензию Creative Commons, если иное не указано в кредитной линии к материалу. Если материал не включен в лицензию Creative Commons, на которую распространяется статья, и его использование не разрешено законом или выходит за рамки разрешенного использования, вам потребуется получить разрешение непосредственно у правообладателя. Чтобы ознакомиться с копией этой лицензии, посетите сайт http://creativecommons.org/licenses/by/4.0/.