
Яркая иллюстрация, позаимствованная из статьи (Zeng M, Gentekaki E, Hyde KD, Zhao Q, Matočec N, Kušan I. Phylogeny and Morphology of Novel Species and New Collections Related to Sarcoscyphaceae (Pezizales, Ascomycota) from Southwestern China and Thailand. Biology. 2023; 12(1):130 https://doi.org/10.3390/biology12010130) представляет каким огромным разнообразием представлено семейство Sarcoscyphaceae на нашей планете.
Здесь вы видите жизненный цикл Cookeina tricholoma. Cookeina - часто встречающийся в тропических и субтропических регионах род семейства Sarcoscyphaceae, приспособлен к произрастанию во влажной и жаркой среде [1, 2]. Среди собранных авторами исследования образцов большое количество видов из тропических регионов относится к роду Cookeina, что указывает на высокую численность и разнообразие этого рода в Китае и Таиланде.
Но вернемся к началу статьи. Вот, что сообщается о семействе, представителей которого мы привыкли наблюдать ранней весной как что-то экзотическое в наших лесах. Далее следуют фрагменты перевода текста статьи, без кавычек, свои пояснений буду выделять шрифтом.
Виды семейства саркосцифовые (Sarcoscyphaceae) являются сапротрофами и растут на опавших ветвях, пнях, стволах или сучьях. Большинство представителей этого семейства широко распространены в тропических районах, и лишь небольшая часть известных видов встречается в районах с умеренным климатом. Все виды имеют типичные плодовые тела в форме диска или чашечки различной окраски - от белого, серого, оранжевого, красного до коричневого. Известно о большом разнообразии саркосцифовых в юго-западном Китае и Таиланде. В этом исследовании приводятся новые описания пяти известных видов и выделяется три новых вида из семейства саркосцифовых из этих регионов на основе морфологии и филогении. Также предлагается поправка для Phillipsia gelatinosa. Впервые о появлении Cookeina sinensis, распространенного в Китае вида, сообщается из Таиланда.
Семейство саркосцифовые (Pezizales) отличается мелкими и крупными, ярко окрашенными сидячими и прилепляющимися апотециями, многоядерными и пигментированными парафизами, опушенными аскоспорами с толстыми стенками и многоядерными, одноядерными и многозубчатыми аскоспорами с гладкими стенками или орнаментом. Авторами было собрано более 40 образцов представителей Sarcoscyphaceae с опавших веток или древесины. Исходя из морфологии и филогении, эти виды относятся к родам Cookeina, Nanoscypha, Phillipsia, Pithya и Sarcoscypha. Среди них представлены три новых вида – Nanoscypha aequispora, Pithya villosa и Sarcoscypha longitudinalis. Филогенетический анализ, основанный на областях генов ITS, LSU, SSU, rpb2 и tef-1α, указывает на родство этих видов внутри семейства саркосцифовых. В то же время было предложено Ph. gelatinosa как синоним Ph. domingensis. Сообщается об одном новом обнаружении C. sinensis в Таиланде.
История исследований
К семейству саркосцифовых относятся дискомицетные грибы, которые в изобилии встречаются в тропических районах, но также встречаются и в регионах с умеренным климатом [3, 4, 5]. Ле Гал [6] неправильно представил семейство саркосцифовых, не предоставив латинского описания. Экблад [7] предоставил достоверное описание в соответствии с надлежащими стандартами номенклатуры. Однако перевод типового рода Sarcosomataceae в Sarcoscyphaceae вызвал долговременную концептуальную путаницу между этими двумя семействами [6, 7, 8, 9]. Только после разъяснения Корфа [10] у Sarcoscyphaceae появилась четкая концепция. Семейство Sarcoscyphaceae имеет типичные апотеции для пецицомицетов (обычно называемых чашевидными грибами) и относится к одному из немногих семейств, в котором отсутствуют сведения о гипогенных таксонах [4]. Апикальный аппарат аска, один из самых характерных признаков всей группы аскомицетов, на протяжении десятилетий вызывал недоумение. Шадефо [11] и Ле Гал [6] рассмотрели некоторые апикальные структуры, которые, по-видимому, были гипотетическими структурами и/или артефактами, возникшими в процессе фиксации материала. Они предположили, что эти структуры представляют собой переход между иноперкулярным апикальным кольцом и настоящей крышечкой. Они назвали этот вид аскуса ‘параоперкулярным’ или ‘субоперкулярным’. Они предположили, что этот аскус представлял собой промежуточную стадию на пути эволюции чашевидных форм.
Экблад [7, 12], ван Бруммелен [13, 14], Самуэльсон [15] и Самуэльсон с соавторами [16] показали, что тип апикального аппарата саркосцифы на самом деле это круглая форма, которая ни в коем случае не является переходной формой между верхушечным кольцом аскуса и настоящей пецициевой головкой. Впоследствии это было подтверждено многочисленными филогенетическими анализами. Sarcoscyphaceae характеризуется ярко окрашенными, от сидячих до прилепившихся апотеций, пигментированными парафизами, содержащими каротиноиды, толстостенными аскоспорами, оснащенными узкой и толстой линзовидной крышечкой, окруженной субапикальной заметно утолщенной стенкой (субоперкулюм), и аскоспорами от одногнездных до многогнездных с гладкими стенками или цианофобными поперечными бороздками/сетчаткой [4, 8]. Имеется всего несколько сообщений об анаморфах в этом семействе. Пфистер [4] и Эканаяка и др. [5] предоставили самые последние обзоры. В семействе насчитывается 13 родов, а именно Aurophora (Rifai), Cookeina (Kuntze), Geodina (Denison), Kompsoscypha (Pfister), Microstoma (Bernstei), Nanoscypha (Denison), Phillipsia (Berk), Pithya (Fuckel), Pseudopithyella (Seaver), Rickiella (Syd. & P. Syd. ex Rick), Sarcoscypha, Thindia (Korf & Waraitch) и Wynnea (Berk. & M.A. Curtis). Всего в этом семействе насчитывается 83 вида [17, 18]. В семействе саркосцифовых несколько видов используются в пищу и медицине. Например, Cookeina speciosa (Деннис) и Cookeina tricholoma (Кунце) считаются съедобными грибами в Мексике, в то время как в Камеруне также есть сведения об их использовании для лечения ушных инфекций [19, 20, 21]. Чашечка алого эльфа, Sarcoscypha coccinea (Ламботт) считается также съедобной [22].
Далее авторы рассказали историю классификации саркосциф исходя из генетических исследований.
Самое раннее филогенетическое исследование Sarcoscyphaceae было проведено Харрингтоном и соавторами [23], которые использовали участок гена малой субъединицы рРНК (SSU) для реконструкции филогении Pezizales. Филогенетический анализ девяти последовательностей, относящихся к девяти родам, показал монофилию Sarcoscyphaceae и его принадлежность к Pezizales [23]. Ромеро и соавторы [17] добавили молекулярные данные по известному виду Rickiella и изучили филогенетические связи внутри Sarcoscyphaceae на основе последовательностей ядерной большой субъединицы рРНК (LSU) и малой субъединицы рДНК. Анджелини и соавторы [24] описали новый вид Geodina на основе морфологии и филогенетического анализа с использованием LSU, но позже Пфистер и соавторы [25] показали, что это синоним типового вида. Пфистер и соавторы [25] предложили новое семейство Wynneaceae, в которое вошли Geodina и Wynnea, тем самым отделив эти таксоны от Sarcoscyphaceae на основании филогенетического анализа четырех генетических маркеров и морфологии. Эти два рода также отличаются морфологическими характеристиками и предпочтениями в среде обитания от других родов Sarcoscyphaceae. Однако участки генов из разных штаммов были объединены для представления определенных таксонов, которые затем использовались для филогенетического анализа. Таким образом, создание нового семейства следует отложить до тех пор, пока не будут получены генетические маркеры интересующих нас таксонов, чтобы избежать путаницы [25]. К сожалению, для Aurophora и Thindia до сих пор нет молекулярных данных, поэтому их филогенетическое положение остается неизвестным.
Не будем вдаваться в подробности методов, как вы знаете, или не знаете, для филогении используется ДНК образца, которая используется для ПЦР на нужный фрагмент гена, который множат для удобства секвенирования его последовательности, а далее полученные коды сравниваются между собой специальными алгоритмами, с помощью программ, в данном случае - DNASTAR Lasergene SeqMan Pro v.7.1.0.
В результате филогенетического анализа, были получены следующие деревья (первое состоит из двух частей, второе получено другим способом, подробности в подписях, специально не буду переводить, чтобы не усложнить понимание терминов):
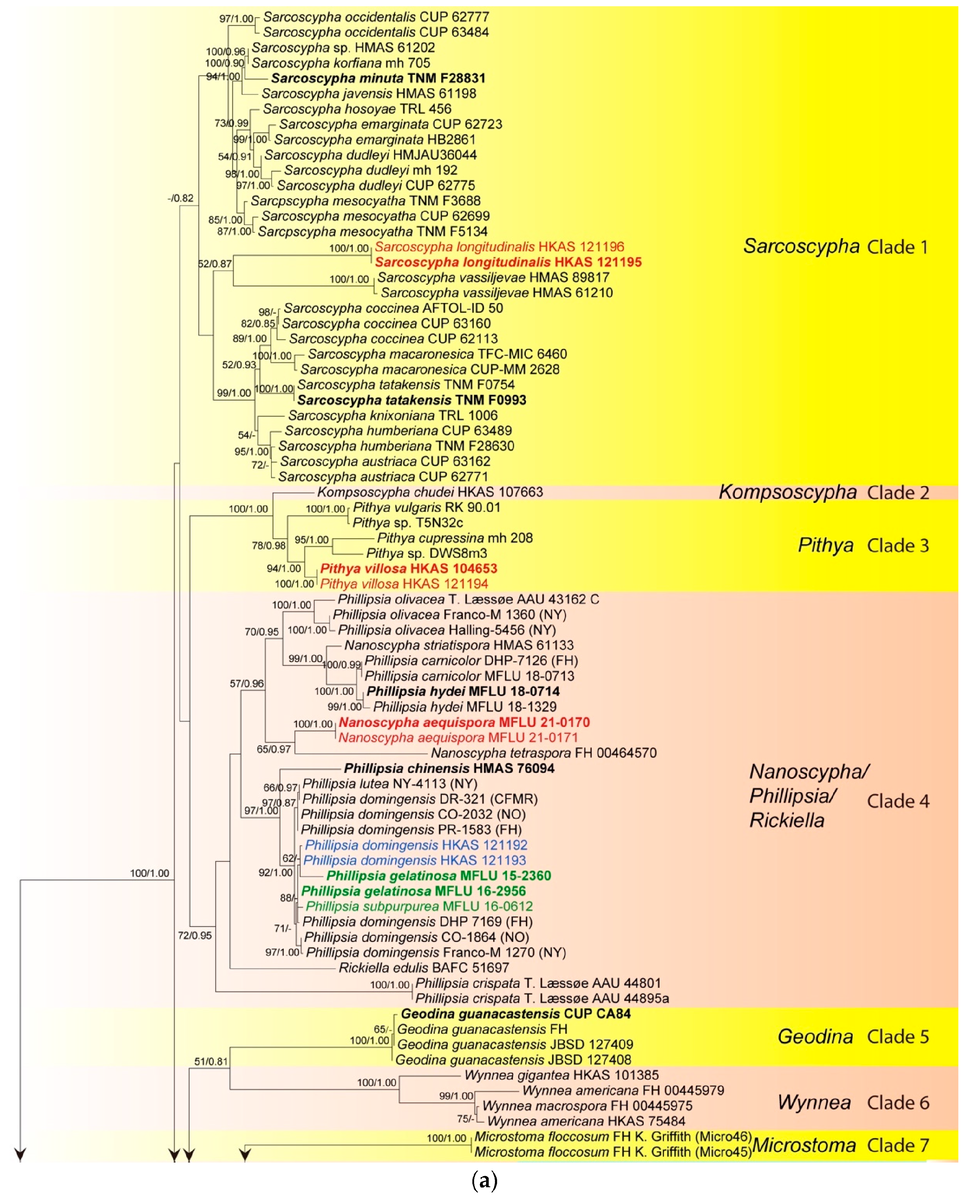
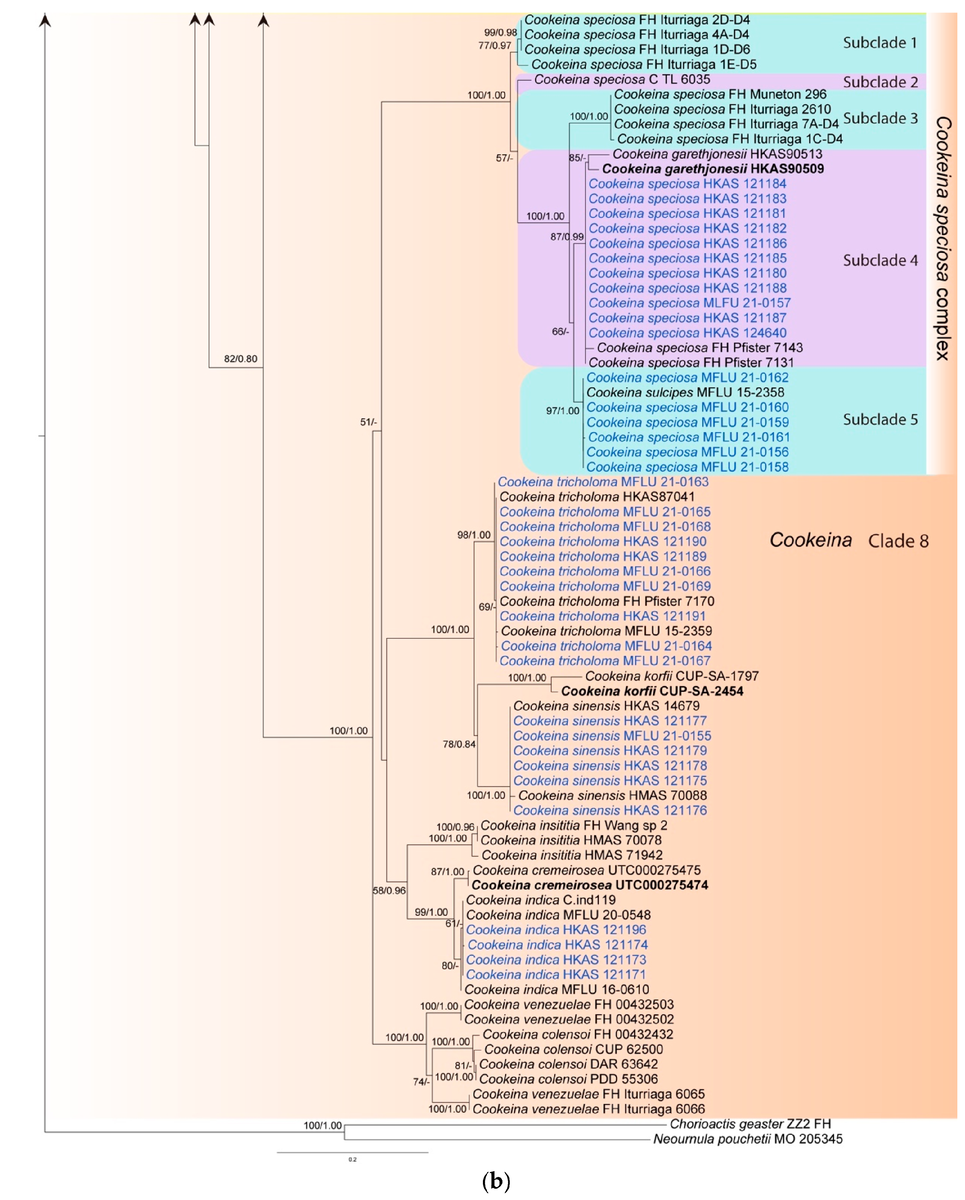
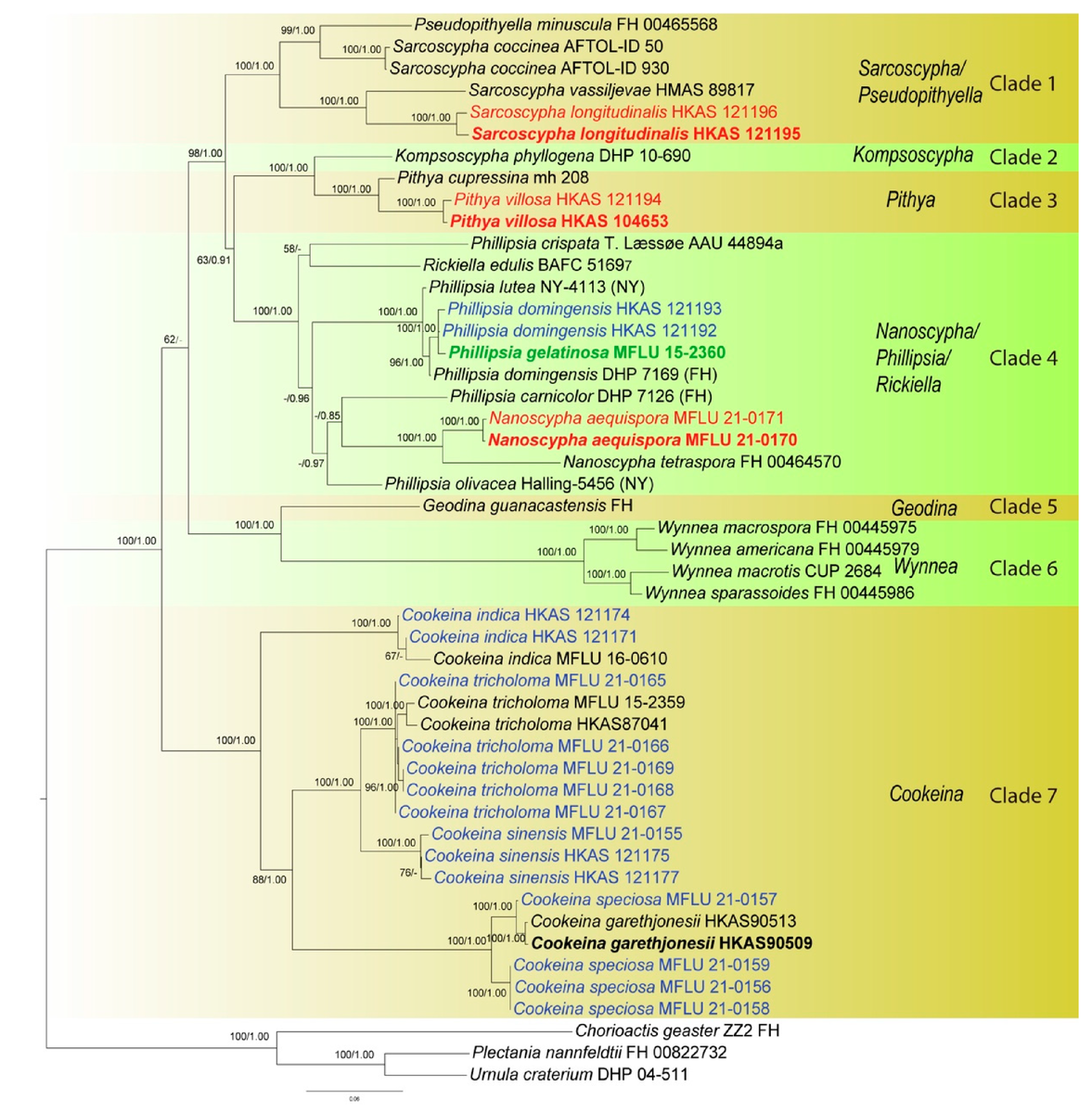
Описание видов, рассмотренных в статье
(Описание неполное, чтобы не перегружать текст терминологией, подробнее в оригинальной статье.)
1. Cookeina indica Pfister & R. Kaushal, Mycotaxon 20(1): 117 (1984)
Растет на опавшей древесине. Телеоморфа: Апотеции высотой до 7 см, шириной 1-4 см, одиночные или рассеянные, глубоко чашевидные, редко в форме колоса, мясистые, с короткой или длинной ножкой. Ножка длиной до 4 см, шириной до 3 мм, от центральной до эксцентрической, заостренная или резко суженная к основанию, с бороздчатым прикреплением, плотная, обычно от белой до желтоватой в свежем виде, желтая в сухом, почти гладкая. Верхняя часть чашеобразной формы, поверхность цвета светлой слоновой кости (RAL 1015), в свежем виде от желтоватой до оранжевой, почти гладкая, с одной концентрической бороздкой, край в целом цельный или редко глубоко раздвоенный с одной стороны. Диск глубоко вогнутый, в основном такого же цвета, как поверхность розетки, или несколько темнее по цвету. Стипальный экто-экзипулум шириной 70–140 мкм, состоит из гиалиновых или желтоватых текстурированных шаровидно-угловатых клеток размером 14–19 × 11–14 мкм. Некоторые наружные шаровидные клетки образуют рыхлые скопления неправильной формы, образующие мучнистую поверхность, иногда с гифоидными волосками шириной 3–6 мкм, гиалиновыми, септатными, слегка сужающимися к закруглённой вершине, обычно собранными в пучки. Стволовая часть состоит из гиалиновых или субгиалиновых текстурированных гиф шириной 3–6,5 мкм. Эктодерма 80-130 мкм толщиной, в основном разделен на два подслоя, ограниченных внешними и внутренними клетками: внешний слой, состоящий из гиалиновой до желтоватой текстуры шаровидной до текстуры призматической, концевые клетки шаровидные, диаметром 13-19 мкм, с пруинозоподобной поверхностью, призматические клетки 23-31 × 11-16 мкм, гиалоидные волоски, образующие по краю обильные пучки, состоящие из субгиалиновых перегородчатых гиф шириной 4-6 мкм, иногда с отдельными монилиоидными волоскообразными отростками, имеющими 2-3 эллипсоидные клетки; внутренний слой, состоящий из гиалиновых от textura angularis до textura epidermoidea, угловые клетки 14-18 × 11-14 мкм и удлиненные клетки 16-23 × 6-9 мкм. Срединная пластинка шириной 200–260 мкм, состоит из гиалиновых текстурированных гиф шириной 4–7 мкм. Гименофор толщиной 340–400 мкм, субгиалиновый, парафизы в обезвоженном состоянии немного выступают за пределы асков. Парафизы шириной 4–6 мкм в средней части, цилиндрические, септатные, суженные в местах перегородок, обычно анастомозирующие, образуя сеть, разветвленные в апикальной части, с заостренной апикальной клеткой. Аски 330–360 × 15–19 мкм, 8-споровые, с эксцентричной крышечкой, цилиндрические, с тупыми верхушками и суженной базальной частью. Аскоспоры [20/1/1, в H2O] (26,3–)29,8–36(–38) × (10,8–)11,5–13,4(–14,2) мкм (Q = 2,08–3,13, Q = 2,64 ± 0,28), веретеновидные или почковидно-веретеновидные, неравносторонние, с небольшими бугорками на полюсах, однорядные, часто с двумя или тремя ямками, иногда с большим количеством ямок, с продольными параллельными гребнями. Анаморф: не обнаружен.
Этот вид отличается почти гладкими апотециями с концентрической бороздкой у края, парафизами с сужающимися концами, веретенообразными и неравносторонними аскоспорами с субпапиллярными концами и продольными бороздками на поверхности аскоспор [26]. Впервые Cookeina indica была обнаружена в Индии, и с тех пор о ней сообщалось в юго-западном Китае [8, 28, 26, 31, 32]. Недавно она также была обнаружена в Таиланде [33]. Этот вид имеет почти гладкую поверхность, если наблюдать его невооруженным глазом, по сравнению с большинством других видов, у которых легко заметны волоски [26], в то время как покрытую мехом поверхность сосуда можно увидеть с помощью ручной лупы. У Cookeina cremeirosea Kropp гладкие апотеции, и она продемонстрировала тесное филогенетическое родство с C. indica (рис. 1) (34). Cookeina cremeirosea отличается розоватыми апотециями и гладкостенными аскоспорами [34]. По сравнению с описанием вида в протологе [31], в представленных коллекциях апотеции крупные (до 7 см в высоту против 3,5 см в высоту) и длинные ножки (до 4 см в длину против 2,2 см в длину). Кроме того, аскоспоры (11,5–13,4 мкм против 10-11,5 мкм) также значительно различаются по ширине.
2. Cookeina sinensis Zheng Wang, Mycotaxon 62: 293 (1997)
Растет на опавшей древесине. Телеоморфа: Апотеции до 4 см высотой, до 5 см шириной, рассеянные, чашевидные, мясистые, с заметной ножкой. Ножка до 2 см длиной, до 8 мм шириной, центральная, черешковая, плотная, в свежем виде от белой до желтоватой, в сухом виде желтая, пушистая или войлочная, с длинными сложными волосками, как на плодоножке. Верхняя часть чашевидная, поверхность от желтоватой до оранжевой или, реже, устрично-белой (RAL 1013) в свежем виде, с длинными сложными волосками, зубчатая, с цельными краями, выровненная или завитковая. Диск глубоко вогнутый, в свежем виде от желтого до оранжевого или розоватого цвета, в основном сливается по цвету с поверхностью плодоножки. Стеблевой внешний слой шириной 90-260 мкм, состоящий из прозрачно-желтоватой шаровидно-угловатой структуры, клеток размером 16-19 × 12-15 мкм, некоторые наружные шаровидные клетки неравномерно, рыхло агрегированы, образуя матовую поверхность. Сердцевина стипаля состоит из гиалиновых или субгиалиновых текстурированных гиф шириной 4–6,5 мкм. Ectal excipulum толщиной 50–110 мкм, состоит из гиалиновой textura globulosa-angularisКлетки размером 15–20 × 12–15 мкм, с гифоидными волосками, образующими многочисленные пучки, состоящие из субгиалиновых, септальных, широких гиф шириной 4–7 мкм, иногда с мониоидоидными отростками, содержащими 1–2 шаровидные клетки, образующие мучнистую поверхность. Медуллярный экципулум толщиной 140–220 мкм, состоит из гиалиновых textura intricata, переплетенных гиф шириной 3–6 мкм. Сложные волоски длиной до 5 мм, диаметром до 150 мкм у основания пучка, пучковидные, от желтого до коричневого цвета, отходят от мозгового вещества экципиума, состоят из параллельных, желтоватых, септатных, толстостенных отдельных волосков диаметром 4–5 мкм с закругленным концом. Гименофор толщиной 280–310 мкм, полупрозрачный. Парафизы шириной 3–4 мкм в середине, нитевидные, с перегородками, анастомозирующие, образующие в средних частях сеть, разветвленные, с закругленным концом. Аски 290–300 × 15–18 мкм, 8-споровые, с эксцентричной крышечкой, цилиндрические, с тупыми верхушками и слегка зауженной базальной частью. Аскоспоры [20/1/1, в H2O] (25–)26.3–29.6(-30.4) × (12.2–)12.4–13.7(-14.2) мкм (Q = 1,91–2,38, Q = 2,15 ± 0,14), веретенообразные, заостренные на концах, односерийные, равносторонние, крупнозубчатые, редко с верхушечковидными затвердениями, присутствующими на одном полюсе, гладкие. Анаморф: не замечен.
У этого вида верхняя часть покрыта длинными сложными волосками, растущими из сердцевинного отростка. Аскоспоры веретенообразные, гладкие, с апикулоподобной структурой [8, 26]. Cookeina sinensis похожа на C. tricholoma и C. korfii Iturr., F. Xu & Pfister, так как у всех трех видов есть заметные длинные сложные волоски. По сравнению с C. korfii и C. sinensis, у C. korfii аскоспоры меньшего размера (22-25 × 9-11,5 мкм) [29]. Основное различие между C. sinensis и C. tricholoma заключается в том, что аскоспоры C. tricholoma имеют продольные бороздки, тогда как у C. sinensis аскоспоры гладкие [26]. О Cookeina sinensis сообщалось только в Китае [6, 26, 30], пока Патил и соавторы [31] не описали новые коллекции из Индии. Сравнивая индийские образцы с представленными здесь, можно отметить, что индийские образцы имеют более крупные аскоспоры (30-40 × 15,6 мкм) [31]. Аскоспоры представленных здесь образцов имеют сходный размер с типовыми образцами (25-28 × 12-12,5 мкм), описанными Вангом [30], у которых апотеции меньшего размера (высотой 2,5 см), чем у здесь. В этом исследовании сообщается о новом исследовании, полученном в Таиланде (MFLU 21-0155).
3. Cookeina speciosa (Fr.) Dennis, Mycotaxon 51: 239 (1994)
Растет на опавшей древесине. Телеоморфа: Апотеции высотой до 4 см, шириной до 3 см, редко до 6 см, одиночные или рассеянные, чашевидные, воронковидной формы, обычно с длинной ножкой. Ножка длиной до 2,5 см, шириной до 3 мм, центральная, черешчатая, твердая, в свежем виде от белой до желтоватой, в сухом виде желтая, пушистая или войлочная. Верхняя часть чашевидной формы, поверхность от желтоватой до оранжевой, бледно-розовой, цвета светлой слоновой кости (RAL 1015) или от розоватой до темно-коралловой, редко белая в свежем виде, с 5 отчетливыми концентрическими гребнями, состоящими из сложных волосков, с цельными краями или слегка завитыми, с длинными волосками. Диск глубоко вогнутый, в свежем виде от желтоватого до оранжевого или от бледно-розового до розового, в основном такого же цвета, как поверхность плодоножки, или более темного цвета. Эктоплазма стебля шириной 50-140 мкм, состоящая из прозрачно-желтоватой текстуры шарообразно-угловатых клеток размером 18-27 × 13-24 мкм, некоторые наружные шаровидные клетки неравномерно, рыхло агрегированы, образуя матовую поверхность. Stipal medulla состоит из гиалиновой и субгиалиновой сложной структуры, гиф шириной 3-7 мкм. Эктальный бугорок толщиной 60-160 мкм, состоящий из гиалиновой шарообразно-угловатой структуры, клеток размером 17-28 × 12-22 мкм, монилиоидных отростков, обычно состоящих из 1-2 округлых клеток, иногда с субклаватидными концевыми клетками, образующими матовую поверхность, сложные пучки волос длиной до 800 мкм и шириной до 260 мкм на концах, основание желтое, состоит из желтоватых перегородчатых гиф шириной 5-8 мкм, которые срастаются в пучки треугольной формы. Склеродерма толщиной 60–250 мкм, состоит из гиалиновых текстурированных гиф, переплетенных между собой, шириной 2–6 мкм. Гименофор толщиной 280–340 мкм, субгиалиновый, с гимениальными щетинками шириной 4–8 мкм, выступающими за пределы гименофора максимум на 45 мкм, с 1–2 перегородками, с закругленной верхушкой. Парафизы шириной 2–4 мкм в средней части, нитевидные, септатные, в основном суженные в местах перегородок, анастомозирующие, образуя сеть, разветвленные, с закругленной верхушкой. Аски 290–320 × 17–21 мкм (подклад 4) или 266–300 × 14–17 мкм (подклад 5), 8-споровые, эксцентрично оперкулюмные, цилиндрические, с тупыми верхушками и узким гифоидным основанием. Аскоспоры [20/1/1, в H2O] (24.8–)25.3–27.5(-28.1) × (13.1–)13.7–15(-15.5) мкм (Q = 1,69–1,96, Q = 1,84 ±0,07) (подкласс 4) или (19.6–)22.2–24.7(-25) × (9.2–)10.3–11.9(-12.3) мкм (Q = 1,93–2,38, Q = 2.12 ± 0,11) (подкласс 5), эллипсоидные, закругленные на концах, однорядные, равносторонние, в зрелом состоянии с двумя ямками, в незрелом — с несколькими ямками, с выступающими апикулюсами на одном или обоих полюсах, периспора с анастомозирующими цианофобными продольными бороздками. Анаморф: не обнаружен.
Этот вид широко распространен в тропических районах [26]. Главной особенностью этого вида является изменчивая окраска апотеция с пятью отчетливыми концентрическими гребнями по краям и сложными волосками, расположенными на этих гребнях. Кроме того, гимений имеет перепончатые щетинки, аскоспоры широкой эллипсоидной формы с явно выступающими верхушками и продольные бороздки на поверхности аскоспор, в некоторых местах соединенные анастомозами [1, 26]. Этот вид был введен Деннисом [27] на основе названия Peziza speciosa Fr.; между тем, C. sulcipes был синонимизирован как более поздний эпитет [27]. Хотя филогенетические исследования, основанные на нем, показывают некоторые генетические вариации, связанные с различиями в окраске внутри клада C. speciosa [1], Итурриага и Пфистер [26] по-прежнему рассматривают это как комплекс. В этом исследовании Cookeina garethjonesii, представленная двумя штаммами, образовала независимую ветвь, которая входит в кладу C. speciosa в подклассе 4 (Fig. 1). Представленная здесь филогения содержит исчерпывающую выборку последовательностей C. speciosa. Cookeina garethjonesii была выделена в отдельный вид от C. speciosa из-за отсутствия перепончатых щетинок и гладкостенных аскоспор [28]. На иллюстрации голотипа C. garethjonesii, представленной в оригинальном исследовании, отчетливо видны отчетливые перепончатые щетинки [28] (Fig. 2 в оригинальной статье). В своей филогении виды были филогенетически различны, однако в наборе данных использовалось лишь ограниченное количество последовательностей C. speciosa. Исходя из этих противоречий, предположено, что в будущем следует пересмотреть типовой образец. На представленных деревьях шесть коллекций были сгруппированы в последовательности, обозначенной как C. sulcipes (MFLU 15-2358), образуя отдельную кладу в комплексе C. speciosa (подкласс 5, Fig. 1). В этой четко определенной группе есть представители, у которых апотеции от розовых до коралловых почти однородны (рис. 9). Однако апотеции кораллового цвета характерны не только для подкласса 5; скорее всего, коллекции с коралловыми апотециями разбросаны по всему комплексу C. speciosa (например, FH Iturriaga 1E-D5, FH Iturriaga 4A-D4 и FH Iturriaga 7A-D4 из подклассов 1 и 3, Fig. 1) [1] (это исследование). В описанных здесь коллекциях C. speciosa (MFLU 21-0157) (Fig. 8) из подкласса 4 имеет более широкие аскоспоры (17-21 мкм против 14-17 мкм) и более крупные аскоспоры (25,3–27,5 × 13,7–15,0 мкм против 22,2–24,7 × 10,3–11,9 мкм) по сравнению с C. speciosa (MFLU 21-0162). (Fig. 10) из подкласса 5. Тем не менее, C. sulcipes (MFLU 15-2358), описанный Эканаякой и соавторами [28], имеет неотличимые размеры аск (280-380 × 15-22 мкм) и аскоспор (21-30 × 11-18 мкм) по сравнению с обеими описанными здесь коллекциями. Следовательно, по-видимому, размеры аск и аскоспор не позволяют отличить эти экземпляры на видовом уровне. В рамках комплекса C. speciosa коллекции авторов разделены на два подкласса, оба из которых имеют высокую статистическую поддержку (подкласс 4: 87BS/0,99п.л., подкласс 5: 97BS/1,00 п.л.). Для выявления комплекса C. speciosa в будущем необходимы дополнительные исследования образцов и видов.
4. Cookeina tricholoma (Mont.) Kuntze, Revis. gen. pl. (Leipzig) 2: 849 (1891)
Растет на опавшей древесине. Телеоморфа: Апотеции высотой до 3 см, шириной до 2 см, одиночные или рассеянные, чашевидные, мясистые, с короткой или длинной ножкой. Ножка до 1,5 см длиной, до 2 мм шириной, центральная, редко эксцентричная, черешковая, плотная, желтоватая или розоватая в свежем виде, желтая в сухом, пушистая или войлочная, с длинными сложными волосками в качестве верхушки аскомицета. Верхняя часть чашевидная, в свежем виде поверхность от желтоватой до оранжевой или розовой, с длинными сложными волосками, зубчатая, с цельным краем, редко с глубоким рассечением с одной стороны, в сухом виде скручивается. Диск глубоко вогнутый, в свежем виде от желтого до оранжевого или розоватого цвета, в основном сливается по цвету с поверхностью плодоножки. Эктоплазма стебля шириной 85-175 мкм, состоящая из прозрачно-желтоватой шарообразно-угловатой структуры, клеток размером 12-17 × 11-15 мкм, некоторые наружные шаровидные клетки неравномерно, рыхло агрегированы, образуя матовую поверхность. Stipal medulla состоит из гиалиновых волокон, гиф шириной 5-7,5 мкм. Эктальный бугорок толщиной 50-80 мкм, состоящий из гиалиновой текстуры шарообразно-угловатой, клетки 13-19 × 11-15 мкм, с двумя типами волосков, смешанных по всей поверхности эктального бугорка: пучковидные гифоидные волоски, состоящие из субгиалиновых и желтоватых перегородчатых широких гиф, шириной 5-9 мкм; дополнительно с монилоидными отростками, состоящими из 2 или более шаровидных отростков, клетки, образующие матовую поверхность. Срединное влагалище шириной 100–160 мкм, состоит из гиалиновых текстурированных гиф, переплетенных между собой, шириной 3–5 мкм. Сложные волоски длиной до 5 мм, диаметром до 200 мкм у основания пучка, пучковидные, коричневые, отходят от срединного влагалища, состоят из параллельных, желтоватых, септатных, толстостенных гиф диаметром 3–6 мкм с закругленным концом. Гименей толщиной 280–350 мкм, субгиалиновый. Парафизы шириной 1,5–3 мкм в средней части, нитевидные, септатные, анастомозирующие, образуя сеть, разветвленные, с закругленным концом. Аски 308–342 × 13–21 мкм, 8-споровые, эксцентрически оперкулюмные, цилиндрические, с тупыми верхушками и узкими основаниями. Аскоспоры [20/1/1, в H2O] (25,8–)28,0–32,9(–33,9) × (11–)11,6–12,8(–13,8) мкм (Q = 2,33–2,68, Q = 2.50 ± 0,17), веретенообразные, заостренные на концах, однорядные, равносторонние, с двумя или более ямками, иногда с выступающими апикулюсами на одном из полюсов, с орнаментом в виде тонких продольных бороздчатых гребней. Анаморф: не обнаружен.
Этот вид отличается апотециями от желтого до оранжевого, или коралловыми, с длинными сложными волосками, отходящими от сердцевинного бугорка, веретенообразными аскоспорами с продольными бороздками [26]. Он похож на C. korfii и C. sinensis, у которых аскоспоры гладкостенные [29]. Этот вид широко распространен в тропических районах, а также является распространенным видом на юго-западе Китая и в Таиланде [28, 26].
5. Nanoscyphaaequispora M. Zeng, Q. Zhao & K.D. Hyde, sp. nov.
Этот вид определяется по раковинным или неглубоко чашевидным апотециям с широкой беловатой ножкой, диску от желтоватого до оранжевого, волнистому краю, нитевидным парафизам с желтоватыми гранулами, эллипсоидным и равносторонним аскоспорам с бигуттулейтом.
Растет на сухой древесине и плодах растений. Телеоморфа: Апотеции высотой 0,5–1 мм, шириной 1-2 мм, рассеянные, в свежем виде с мелкими чашелистиками, в сухом - с раковиной, с широкими прилистниками, голые. Ножка длиной 400-1500 мкм, шириной 500-2000 мкм, центральная, воронкообразная, морщинистая на поверхности, плотная, от беловатого до кремового, реже желтоватого цвета. Плодоножка неглубоко-чашевидная, поверхность от желтоватой до оранжевой, края волнистые. Диск от неглубоко-вогнутого до дисковидного, одного цвета с поверхностью плодоножки. Стеблевой эктоэкципулят 62-166 мкм, состоящий из гиалиновой структуры angularis, клетки 14-20 × 8-12 мкм, смешанной с призматической структурой, клетки 18-25 × 9-12 мкм, с несколькими порректоидными клетками, расположенными на поверхности, клетки шириной 4-6 мкм. Stipal medulla состоит из гиалиновых волокон, гиф шириной 4-6 мкм. Эктальный эксципулюм толщиной 56-94 мкм, состоящий из гиалиново-желтоватой глобулезно-угловатой текстуры, клетки 14-22 × 9-13 мкм, смешанной с призматической текстурой, клетки 19-25 × 9-13 мкм, с несколькими порректоидными клетками, расположенными на поверхности, клетки шириной 4-6 мкм. Слой septate hyphae толщиной 76-192 мкм, состоящий из гиалиновой структуры, гиф шириной 4-6 мкм. Гимений толщиной 280-310 мкм, желтоватый, парафизы при обезвоживании немного превышают размеры asci. Парафизы шириной 2-3 мкм в средней части, нитевидные, разветвленные, перегородчатые, заполненные желтоватыми гранулами. Чешуйки 235-284 × 10-13 мкм, 8-споровые, нижнечелюстные, верхушки тупые, цилиндрические, сужающиеся к основанию. Аскоспоры [20/1/1, в H2O] (14.8–)16.2–18.6(-19.2) × (12.4–)10.3–11.6(-9.9) мкм (Q = 1,41–1,78, Q = 1,59 ± 0,14), эллипсоидная, с округлыми или слегка усеченными концами, равносторонняя, реже слегка неравносторонняя с одной стороны плоские, однорядные, в незрелом состоянии многозубчатые, в зрелом - крупнозубчатые, с гладкими стенками. Анаморф: не замечен.
Виды рода Nanoscypha - это небольшие чашевидные грибы, обычно менее 1 см в диаметре. Апотеции варьируют от дисковидных до чашевидных, от раковинных до воронкообразных, от сидячих до прилистниковых, от желтого до оранжевого и красного цвета. Цилиндрические аскоспоры с 3-, 4-, 6- или 8-ми спорами и заостренными основаниями. Аскоспоры от эллипсоидных до почковидных, в основном неравносторонние, редко равносторонние, в основном с двумя масляными каплями [8, 35]. В настоящее время к этому роду относят восемь видов в соответствии с Species Fungorum [36], в то время как размещение N. striatispora (W.Y. Zhuang) F.A. Harr. [8, 35] все еще обсуждается (см. обсуждение). Хотя этот новый вид и N. tetraspora объединены в одну группу, трудно подтвердить правильное положение нового вида на филогенетическом древе из-за отсутствия доступных данных по другим видам. Между тем, штаммы Nanoscypha включены в кладу вместе с Phillipsia и Rickiella в ITS и мультигенном анализах. Благодаря морфологическому сравнению, большинство видов имеют неравносторонние аскоспоры, за исключением N. macrospora Denison [8, 37, 38] и двух неопределенно описанных видов, N. bella (Berk. & M.A. Curtis) Pfister и N. euspora (Rick), т. е. карп. Из этих трех видов у N. bella апотеции и аскоспоры более крупные [39], в то время как N. euspora отличается своими аскоспорами, не имеющими зубцов [40]. У Nanoscypha macrospora равносторонние аскоспоры, реже неравносторонние, как у этого нового вида. Основное отличие от N. macrospora заключается в том, что аскоспоры содержат только 3 или 4 аскоспоры. Кроме того, аскоспоры имеют вытянутую эллипсоидную форму [35]. Более того, N. orissaensis C.M. Das & DC. Pant - редко регистрируемый вид, для которого не хватает типового материала [38]. Таким образом, мы предложили новый вид Nanoscypha aequispora, основанный на морфологии.
6. Phillipsia domingensis (Berk.) Berk. ex Denison, Mycologia 61(2): 293 (1969)
Растет на опавшей древесине. Телеоморфа: Апотеции высотой 7-11 мм, шириной до 4 см, разбросанные, кожистые, от неглубоко-чашевидных до дисковидных, с нижними или короткими прилистниками. Ножка длиной до 3 мм, шириной до 5 мм, от центральной до эксцентричной, конической формы, цельная, ярко-бежево-красная (RAL 3012), или красноватая, или кремово-желтоватая, опушенная. Верхушка неглубоко чашевидная, поверхность одного цвета с ножкой, опушенная, края цельные. Диск от неглубокой чашеобразной до дисковидной формы, жемчужно-розовый (RAL 3033) или темно-красный (RAL 3031) или с желтыми вкраплениями. Стеблевая эктоплазма толщиной 60-90 мкм, состоящая из желтоватой или субгиалинизированной структуры, от прямой до эпидермоидной, гиф шириной 4-7 мкм, с некоторыми внешними гифами, которые неравномерно и рыхло расположены, образуя опушенную поверхность. Stipal medulla состоит из гиалиновых волокон, гиф шириной 3,5–5 мкм. Эктальный бугорок толщиной 60-100 мкм, состоящий из желтоватой или субгиалинизированной структуры, от прямой до эпидермоидной, гиф шириной 4-7 мкм, с несколькими рыхлыми гифами в самой внешней части. Оболочка septate hyphae толщиной 280-650 мкм, состоящая из гиалиновых волокон, гиф шириной 3-4 мкм. Гимений толщиной 300-350 мкм, от розового до красного, парафизы при обезвоживании немного превышают размеры aск. Парафизы шириной 1–2 мкм в средней части, нитевидные, с красноватым содержимым, септатные, разветвленные. Аски 285–346 × 11–14 мкм, 8-споровые, с эксцентричной крышечкой, цилиндрические, с тупыми верхушками, сужающиеся к основанию. Аскоспоры [20/1/1, в H2O] (19.8–)20.5–24.2(-27.7) × (10.4–)10.6–12(-13.2) мкм (Q = 1,82–2,17, Q = 1,98 ± 0,08), подчелюстная или почковидная форма с заостренными или субпапиллярными концами, односерийная, неравнобоковая, однозубчатая или крупнозубчатая, украшенная несколькими продольными бороздками. Анаморф: не замечен.
Это распространенный вид в субтропических и тропических районах. У этого вида апотеции более крупного размера, гимений от красного до пурпурно-красного, субреновидные или почковидные аскоспоры с несколькими заметными продольными бороздками [58, 59]. Хансен и др. [60] предположили, что комплекс Ph. domingensis основан на генетическом маркере ITS, благодаря виду Ph. lutea Denison и некоторым коллекциям Ph. domingensis с желтыми апотециями, которые гнездились в типично красном Ph. domingensis. Эканаяка и др. [61] представили Ph. gelatinosa на основе трех коллекций и представили описание Ph. subpurpurea Berk. И Брум вместе с молекулярными данными, которых раньше не существовало. Даже эти два вида включены в комплекс Ph. domingensis, Ekanayaka и др. [61] выявили морфологические различия, позволяющие отличить их от Ph. domingensis. Phillipsia gelatinosa отличается оранжевым содержанием парафиз, аски и аскоспор большего размера и наличием студенистой оболочки, окружающей аскоспоры [61]. Phillipsia subpurpurea (MFLU16-0612) отличается тем, что имеет гладкие аскоспоры или со слабыми бороздками, толстую студенистую оболочку, окружающую аскоспоры. Мы повторно исследовали все образцы, названные Ph. gelatinosa и Ph. subpurpurea (MFLU16-0612) (Fig. 15, 16 и 17). Три из них обладают морфологическими признаками, почти соответствующими Ph. domingensis, которые характеризуются отчетливым красноватым содержимым парафизов, субреновидных или почковидных аскоспор с заметными продольными бороздками. Хотя у Ph. gelatinosa (MFLU 15-2360) трудно обнаружить заметное красноватое содержимое и полосатые аскоспоры из-за качества образца, мы все же можем обнаружить спорадическое красноватое содержимое в парафизах и слабые бороздки на поверхности аскоспор. У всех образцов структура, напоминающая оболочку, присутствовала только при повторном увлажнении в 10% растворе КОН, но не в воде. Сравнивая размеры аск и аскоспор повторно исследованных образцов, мы не обнаружили существенных различий между этими образцами, в то время как эти размеры в значительной степени отличаются от описаний Ekanayaka et al. Самое главное, что “два голотипа” (MFLU 16-2956 и MFLU 15-2360) были отнесены к протологу Ph. gelatinosa [61]. В соответствии с Международным кодексом номенклатуры водорослей, грибов и растений, в частности, статьей 8.1 Кодекса Шэньчжэня [62], Ph. gelatinosa - неправильное название. Исходя из этого, Ph. gelatinosa должен быть недействительным наименованием и синонимом Ph. domingensis. Между тем, образец (MFLU16-0612) был неправильно отнесен к Ph. subpurpurea, что следует исправить на Ph. domingensis. Phillipsia domingensis - сложный вид, у которого отсутствуют последовательности типов, и для большинства коллекций известен почти только его регион. Таким образом, для эффективной дифференциации на видовом уровне необходимо секвенирование дополнительных участков ДНК и получение большего количества данных о фенотипических признаках как можно большего числа коллекций.
7. Pithya villosa M. Zeng, Q. Zhao & K.D. Hyde, sp. nov.
Этот вид определяется по мелким чашевидным или дисковидным или выпуклым апотециям, растущим на Juniperus sp. (можжевельник), желтоватой поверхности, покрытой гифовидными волосками, цельнокрайнему или лопастному краю, субхиалиновым или желтоватым парафизам, сферическим аскоспорам с зернистым содержимым.
Сапробен на веточках Juniperus sp. Телеоморфа: Апотеции высотой 2-3 мм, шириной 3-6 мм, от рассеянных до групповых, мясистые, от неглубоко-чашевидных до дисковидных или выпуклых, от сидячих до замещающих. Верхняя часть неглубоко чашевидная, в свежем виде цельное до лопастевидной, в сухом - скрученная, от полугладкой до мелкоопушенной, беловатой, с боков от опушенной до ворсинчатой к основанию, беловатая на желтоватой поверхности. Диск от дисковидного до слегка выпуклого, от желтого до оранжевого. Эктальный бугорок шириной 60-100 мкм, гиалиновый на более широкой краевой поверхности, состоит из пористой структуры, от субгиалиновой до желтоватой к основанию, и состоит из эпидермоидной структуры с клетками шириной 5-8 мкм, до угловатой структуры с клетками размером 13-17 × 9-12 мкм. Волоски в основном растут на боковой поверхности эксципулеза и апотециальном основании, субгиалиновые, изгибающиеся, гифоидные, септированные, шириной 5-7 мкм. Сердцевинная оболочка шириной 60-210 мкм, сложной структуры, гиалиновая, состоит из гиф шириной 4-6 мкм. Гимений толщиной 180-290 мкм, желтый, парафизы при обезвоживании немного превышают аск. Парафизы шириной 2-3 мкм в средней части, нитевидные, на вершине увеличенные, шириной 4-6 мкм, разветвленные, септированные, от субхиалинового до желтоватого цвета. Чешуйки 227-275 × 11-14 мкм, 8-споровые, конечные, субцилиндрические, на вершине тупые, сужающиеся к основанию. Аскоспоры [20/1/1, в H2O] (11.4–)11.7–13.7(-14.4) × (11.3–)11.7–13.8(-14.0) мкм (Q = 0,93–1,14, Q = 1,00 ± 0,05), сферическая, однорядная, субгиалиновая, с преломляющим зернистым содержимым, гладкостенная. Анаморф: не замечен.
Пития уникальна среди саркосцифовых своими сферическими аскоспорами [35]. Большинство видов имеют сходные признаки, а именно апотеции от желтого до оранжевого, от неглубоких чашевидных до дисковидных, от сидячих до субстипитных, нитевидные парафизы с увеличенными верхушками и гладкостенные сферические аскоспоры [35, 42, 43]. Kirk et al. [44] и Wijayawardene et al. [18, 45], соответственно, отнесли к этому роду пять и два вида без указания видовых названий, в то время как в Species Fungorum имеется 11 записей о видах [36], исключая Pi. thujina (Peck) Sacc. который является синонимом Pi. cupressina [42]. Насколько нам известно, видовое богатство этого рода, по-видимому, не выяснено. Введение нового вида основано на сравнении десяти видов Pithya в соответствии с данными Species Fungorum. Pithya cupressina и Pi. vulgaris Fuckel - два древнейших вида, интродуцированных в этот род, и они также наиболее часто регистрируются [35, 42, 43, 45, 46-52]. Молекулярные данные также наиболее многочисленны по этим двум видам [17, 23, 53]. У Pithya vulgaris апотеции и аскоспоры крупнее, чем у Pi. cupressina, и в то время как у Pi. обыкновенный в основном растет на отмерших ветвях пихты или сосны обыкновенной, Pi. cupressina обычно растет на кипарисе или можжевельнике обыкновенном [42, 49, 50]. Хотя этот вид также обнаруживается на можжевельнике, он отличается от Pi. cupressina своим верхушечным покровом. В то время как этот новый вид имеет ворсинчатую поверхность, где гифоидные волоски покрывают всю эксципулярную поверхность с краями, и в некоторой степени также более широкую краевую область, Pi. cupressina имеет гладкую эксципулярную поверхность, а белые гифозные волоски присутствуют только у основания [8, 35, 42, 49]. Новый вид отличается филогенетически и является родственным Pi. cupressina. Что касается других видов, которые редко описываются, то большинство из них собраны из сосновых с ограниченным количеством описаний, таких как Pi. arctica Л.И. Васильевой [54], Pi. epichrysea (Beck) Boud. [55], Pi. lacunosa (Ellis & Everh.) Сивер [42], Pi. malochi Velen. и Pi. microspora Velen. [96]. Pi. arethusa Velen. был собран с Ligustrum vulgare (бирючина обыкновенная, маслиновые) [56], а Pi. madothecae Buchloh был собран с Porella platyphylla [57]. Pithya fascicularis (Berk. & Broome) Sacc. был описан только по коре, но его субшаровидных и небольших по размеру аскоспозов (диаметр 7-8 мкм) достаточно, чтобы отличить его от этого нового вида [55].
8. Sarcoscypha longitudinalis M. Zeng, Q. Zhao & K.D. Hyde, sp. nov.
Этот вид можно распознать по коричневым прилистным апотециям с ворсинчатой верхушкой, слегка волнистой или глубоко расщепленной с одной стороны краям, широкими веретенообразными аскоспорам с продольными бороздками.
Растет на опавшей древесине. Телеоморфа: Апотеции до 4 см высотой, 2,5 см шириной, одиночные, чашевидные, прилистные. Ножка длиной до 1 см, шириной 5 мм, центральная, черешковая, плотная, коричневая, с тонкой незаметной подушечкой у основания. Плодоножка чашевидная, ворсинчатая, поверхность коричневая, края слегка волнистые или глубоко раздвоенные с одной стороны. Диск глубоко вогнутый, по цвету совпадает с поверхностью емкости. Прилистник экто-эксципулум шириной 40-80 мкм, состоит из субгиалинового или желтовато-охристого цвета, текстура прямая, гифы шириной 5-7 мкм, у основания обильные гифоидные волоски шириной 4-6 мкм, желтовато-охристые. Stipal medulla состоит из гиалиновой и субгиалиновой сложной структуры, гиф шириной 3,5–6 мкм. Эктальный бугорок шириной 60-100 мкм, состоящий из гиалиновых волокон от прямой до призматической структуры, коричневатый в самой внешней части, гиф шириной 4-7 мкм, с примесью клеток размером 11-16 × 6-8 мкм, с гифоидными волосками на поверхности, шириной 5-7 мкм, перегородчатый, от гиалинового до субгиалинового, с закругленным концом. Сердцевинная оболочка шириной 235-310 мкм, состоящая из гиалиновой и субгиалиновой сложной структуры, гиф шириной 3-4 мкм. Гимений толщиной 280-345 мкм, субгиалинизированный, парафизы при обезвоживании немного превышают аск. Парафизы шириной 2-4 мкм в средней части, нитевидные, разветвленные, перегородчатые, с закругленным концом. Чешуйки 297-359 × 12-14 мкм, 8-споровые, конечные, субцилиндрические, на вершине тупые, сужающиеся к основанию. Аскоспоры [20/1/1, в H2O] (18.3–)19.3–21.4(-22.4) × (9.5–)10.7–12.1(-12.8) мкм (Q = 1,57–1,99, Q = 1,79 ± 0,12), широко-веретенообразные, равносторонние, однорядные, в незрелом состоянии с множеством зубцов, в зрелом - без зубцов, орнамент с несколько продольных бороздок. Анаморф: не замечен.
Саркосцифа отличается серо-белыми, желтыми, оранжевыми или красными апотециями, которые бывают от прилистниковых до прилистничных, от голых до зубчатых, от эллипсоидных до субцилиндрических аскоспор, обычно с тупыми концами или неглубокими углублениями на обоих полюсах, гладкими или орнаментированными стенками, от одногнездных до многогнездных [3, 8, 41, 35]. В семействе Sarcoscypha этот новый вид легко отличить по коричневым прилистным апотециям с ворсинчатой верхушкой, широко-веретенообразным аскоспорам с неигутковидными продольными бороздками. Согласно филогенетическим анализам, этот новый вид является родственным группе S. vassiljevae. Эти два вида имеют сходную морфологию аскоспор, имеющих большую масляную каплю, в то время как S. vassiljevae отличается от нового вида серо-белым гимением и эллипсоидными аскоспорами с гладкими стенками [8].
Обсуждение
Эканаяка и др. [28, 61] предположили, что наличие аскоспоровой оболочки является новым таксономически важным признаком для некоторых новых и известных видов Sarcoscyphaceae. При изучении наших последних коллекций и гербарных образцов эта особенность проявилась только у аскоспор, которые были обработаны 5%-ным или 10%-ным раствором КОН, но не у тех, которые были помещены в воду. Эта ситуация была ранее описана Pfister и соавторами [25] как разрыхление стенок аскоспор при обработке KOH, и, по-видимому, это универсальная характеристика для Sarcoscyphaceae. Таким образом, желатиновая оболочка является недопустимым признаком для описания видов, а тем более подходящим диагностическим признаком для идентификации видов, поскольку она является результатом химической обработки KOH.
В качестве заключения
Юго-Западный Китай и Таиланд являются регионами с высоким видовым богатством семейства саркосцифовых. Виды Cookeina, Phillipsia, и Sarcoscypha очень распространены в этих районах, в то время как данные о Nanoscypha и Pithya ограничены. В настоящем исследовании переописано пять известных видов и выделего три новых вида в эти роды. Между тем, Ph. gelatinosa здесь предлагается в качестве более позднего эпитета Ph. domingensis. Представленные в статье морфологические и филогенетические исследования вносят значительный вклад в продвижение этого семейства к естественной классификации. Однако отсутствие данных о некоторых типовых видах и молекулах, а также наличие комплексов некоторых видов затрудняют будущие исследования.
Список литературы (оригинальная нумерация публикаций изменена под обзор):
1. Weinstein, R.N.; Pfister, D.H.; Iturriaga, T. A phylogenetic study of the genus Cookeina. Mycologia 2002, 94, 673–682.
2. Pfister, D.H.; Kaushal, R. Cookeina indica, a new species from India with a key to the species of Cookeina. Mycotaxon 1984, 20, 117–121.
3. Baral, H.O. Taxonomische und ökologische Studien über Sarcoscypha coccinea agg., Zinnoberrote Kelchbecherlinge. Z. Mykol. 1984, 50, 117–145.
4. Pfister, D.H. Chapter 2. Pezizomycotina: Pezizomycetes, Orbiliomycetes. In Systematics and Evolution. The Mycota (a Comprehensive Treatise on Fungi as Experimental Systems for Basic and Applied Research; McLaughlin, D., Spatafora, J., Eds.; Springer: Berlin, Germany, 2015; Volume 7B, pp. 35–56.
5. Ekanayaka, A.H.; Hyde, K.D.; Jones, E.B.G.; Zhao, Q. Taxonomy and phylogeny of operculate discomycetes: Pezizomycetes. Fungal Divers. 2018, 90, 161–243.
6. Le Gal, M. Les Discomycètes suboperculés. Bull. Trimest. Soc. Mycol. Fr. 1946, 62, 218–240.
7. Eckblad, F.E. The genera of the operculate discomycetes. A re-evaluation of their taxonomy, phylogeny and nomenclature. Nytt Mag. Bot. 1968, 15, 1–191.
8. Zhuang, W.Y. Hyaloscyphaceae, Sarcoscyphaceae et Sarcosomataceae. In Flora Fungorum Sinicorum; Science Press: Beijing, China, 2004; Volume 21, pp. 1–192.
9. Zeng, M.; Gentekaki, E.; Hyde, K.D.; Zhao, Q. Donadinia echinacea and Plectania sichuanensis, two novel species of Sarcosomataceae from southwestern China. Phytotaxa 2021, 508, 1–21.
10. Korf, R.P. Nomenclatural notes. VII. Family and tribe names in the Sarcoscyphineae (Discomycetes) and a new taxonomic disposition of the genera. Taxon 1970, 19, 782–786.
11. Chadefaud, M. Les asques para-operculés et la position systématique de la Pézize Sarcoscypha coccinea Fries ex Jacquin. C R Hebd. Seances Acad. Sci. 1946, 222, 753–755.
12. Eckblad, F.E. The suboperculate ascus, a review. Persoonia 1972, 6, 439–443.
13. van Brummelen, J. Light and electron microscopic studies of the ascus top in Sarcoscypha coccinea. Persoonia 1975, 8, 259–271.
14. van Brummelen, J. The operculate ascus and allied forms. Persoonia 1978, 10, 113–128.
15. Samuelson, D.A. The apical apparatus of the suboperculate ascus. Can. J. Bot. 1975, 53, 2660–2679.
16. Samuelson, D.A.; Benny, L.; Kimbrough, J.W. Asci of the Pezizales. VII. The apical apparatus of Galiella rufa and Sarcosoma globosum: Reevaluation of the suboperculate ascus. Can. J. Bot. 1980, 58, 1235–1243.
17. Romero, A.I.; Robledo, G.; LoBuglio, K.F.; Pfister, D.H. Rickiella edulis and its phylogenetic relationships within Sarcoscyphaceae. Kurtziana 2012, 37, 79–89.
18. Wijayawardene, N.N.; Hyde, K.D.; Al-Ani, L.K.T.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.C.; Zhao, R.L.; Aptroot, A.; Leontyev, D.V.; Saxena, R.K.; et al. Outline of Fungi and fungus-like taxa. Mycosphere 2020, 11, 1060–1456.
19. Villarreal, L.; Perez-Moreno, J. Los hongos comestibles silvestres de Mexico, un enfoque integral. Micol. Neotrop. Apl. 1989, 2, 77–114.
20. Sánchez, J.E.; Martin, A.M.; Sánchez, A.D. Evaluation of Cookeina sulcipes as an edible mushroom: Determination of its biomass composition. In Developments in Food Science; Elsevier: Amsterdam, The Netherlands, 1995; Volume 37, pp. 1165–1172.
21. van Dijk, H.; Onguene, N.A.; Kuyper, T.W. Knowledge and utilization of edible mushrooms by local populations of the rain forest of south Cameroon. Ambio. 2003, 32, 19–23.
22. Arora, D. Mushrooms Demystified: A Comprehensive Guide to the Fleshy Fungi; Ten Speed Press: Berkeley, CA, USA, 1986; pp. 1–836.
23. Harrington, F.A.; Pfister, D.H.; Potter, D.; Donoghue, M.J. Phylogenetic studies within the Pezizales. I. 18S rRNA sequence data and classification. Mycologia 1999, 91, 41–45.
24. Angelini, C.; Medardi, G.; Alvarado, P. Contribution to the study of neotropical discomycetes: A new species of the genus Geodina (Geodina salmonicolor sp. nov.) from the Dominican Republic. Mycosphere 2018, 9, 169–177.
25. Pfister, D.H.; Quijada, L.; LoBuglio, K.F. Geodina (Pezizomycetes: Wynneaceae) has a single widespread species in tropical America. Fungal Syst. Evol. 2020, 5, 131–138.
26. Iturriaga, T.; Pfister, D.H. A monograph of the genus Cookeina (Ascomycota, Pezizales, Sarcoscyphaceae). Mycotaxon 2006, 95, 137–180.
27. Dennis, R.W.G. Plumier’s Discomycetes. Mycotaxon 1994, 51, 237–239.
28. Ekanayaka, A.H.; Hyde, K.D.; Zhao, Q. The genus Cookeina. Mycosphere 2016, 7, 1399–1413.
29. Iturriaga, T.; Xu, F.; Pfister, D.H. Cookeina korfii, a new species hidden in Cookeina tricholoma. Ascomyceteorg. 2015, 7, 331–335.
30. Wang, Z. Taxonomy of Cookeina in China. Mycotaxon 1997, 62, 289–298.
31. Pfister, D.H.; Kaushal, R. Cookeina indica, a new species from India with a key to the species of Cookeina. Mycotaxon 1984, 20, 117–121.
32. Yang, Z.L. Several noteworthy higher fungi from southern Yunnan, China. Mycotaxon 1990, 38, 407–416.
33. Chethana, K.W.T.; Niranjan, M.; Dong, W.; Samarakoon, M.C.; Bao, D.; Calabon, M.S.; Chaiwan, N.; Chuankid, B.; Dayarathne, M.C.; de Silva, N.I.; et al. AJOM new records and collections of fungi: 101–150. Asian J. Mycol. 2021, 4, 113–260.
34. Kropp, B.R. Cookeina cremeirosea, a new species of cup fungus from the South Pacific. Mycoscience 2016, 58, 40–44.
35. Denison, W.C. Central American Pezizales. IV. The genera Sarcoscypha, Pithya, and Nanoscypha. Mycologia 1972, 64, 609–623.
36. Index Fungorum. Available online: http://www.indexfungorum.org/names/names.asp (accessed on 8 September 2022).
37. Harrington, F.A. Relationships among Sarcoscypha species: Evidence from molecular and morphological characters. Mycologia 1998, 90, 235–243.
38. Pant, D.C.; Prasad, V. Indian Sarcoscyphaceous Fungi. In Technology & Engineering; Scientific Publishers: Jodhpur, India, 2008; pp. 1–124.
39. Berkeley, M.J. On a collection of fungi from Cuba. Part II. Bot. J. Linn. Soc. 1869, 10, 341–392.
40. Rick, J.E. Pilze aus Rio Grande do Sul (Brazilien). Brotéria 1906, 5, 5–53.
41. Wang, Y.Z.; Huang, C.L.; Wei, J.L. Two new species of Sarcoscypha (Sarcosyphaceae, Pezizales) from Taiwan. Phytotaxa 2016, 245, 169–177.
42. Seaver, F.J. The North American Cup-Fungi (Operculates); Lancaster Press: New York, NY, USA, 1928; p. 533.
43. Spooner, B. The larger cup fungi in Britain, part 4, Sarcoscyphaceae and Sarcosomataceae. Field Mycol. 2002, 3, 9–14.
44. Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Ainsworth & Bisby’s Dictionary of the Fungi, 10th ed.; CAB International: Wallingford, UK, 2008.
45. Wijayawardene, N.N.; Hyde, K.D.; Rajeshkumar, K.C.; Hawksworth, D.L.; Madrid, H.; Kirk, P.M.; Braun, U.; Singh, R.V.; Crous, P.W.; Kukwa, M.; et al. Notes for genera: Ascomycota. Fungal Divers. 2017, 86, 1–594.
46. Krieglsteiner, G.J. On some new, rare and critical Macromycetes in the Federal Republic of Germany. Z. Mykol. 1985, 51, 85–130.
47. Meléndez-Howell, L.M.; Coute, A.; Mascarell, G.; Bellemère, A. Ultrastructure des asques et des ascospores de Desmazierella acicola (Sarcoscyphaceae, Pezizales, Ascomycetes). Intérêt Systematique et Biologique. Mycotaxon 1998, 68, 53–74.
48. Castellano, M.A.; Smith, J.E.; O’Dell, T.; Cázares, E.; Nugent, S. Handbook to Strategy 1 Fungal Taxa from the Northwest Forest Plan; U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1999; pp. 1–195.
49. Benkert, D. Pithya cupressina und P. vulgaris (Pezizales)–identisch oder nicht? Mycol. Bavarica. 2008, 10, 55–62.
50. Sammut, C. Pithya cupressina (Ascomycota: Pezizomycetes Sarcoscyphaceae): A new addition to the Maltese mycobiota. Cent. Mediterr. Nat. 2012, 5, 54–55.
51. Kunca, V. Pithya vulgaris znovuobjavená na Slovensku. Catathelasma 2015, 16, 11–13.
52. Ortega-López, I.; Valenzuela, R.; Gay-González, A.D.; Lara-Chávez, M.B.N.; López-Villegasy, E.O.; Raymundo, T. La Familia Sarcoscyphaceae (Pezizales, Ascomycota) en México. Acta Bot. Mex. 2019, 126, e1430.
53. Harrington, F.A.; Potter, D. Phylogenetic relationships within Sarcoscypha based upon nucleotide sequences of the internal transcribed spacer of nuclear ribosomal DNA. Mycologia 1997, 89, 258–267.
54. Index of Fungi 2: 444. Available online: http://sftp.kew.org/pub/data-repositories/LibriFungorum/IXF2/IXF2-444.jpg (accessed on 8 September 2022).
55. Saccardo, P.A. Discomyceteae et Phymatosphaeriaceae. Syll. Fung. 1889, 8, 1–1143.
56. Petrak’s Lists 7: 982. Available online: http://sftp.kew.org/pub/data-repositories/LibriFungorum/Petrak7/Petrak7-982.jpg (accessed on 8 September 2022).
57. Döbbeler, P. Octosporella erythrostigma (Pezizales) and Pithyella frullaniae (Helotiales), two remarkable ascomycetes on Frullania dilatate. Feddes Repert. 2004, 115, 5–14.
58. Zhuang, W.Y. Re-dispositions of Phillipsia (Pezizales) collections from China. Mycotaxon 2003, 86, 291–301.
59. Denison, W.C. Central American Pezizales. III. The genus Phillipsia. Mycologia 1969, 61, 289–304.
60. Hansen, K.; Pfister, D.H.; Hibbett, D.S. Phylogenetic relationships among species of Phillipsia inferred from molecular and morphological data. Mycologia 1999, 91, 299–314.
61. Ekanayaka, A.H.; Bhat, D.J.; Hyde, K.D.; Jones, E.B.G.; Zhao, Q. The genus Phillipsia from China and Thailand. Phytotaxa 2017, 316, 138–148.
62. Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.H.; Li, D.Z.; Marhold, K.; et al. International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code) Adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017; Regnum Vegetabile 159; Koeltz Botanical Books: Glashütten, Germany, 2018.