
Глобальное распространение клещей Varroa destructor — одно из самых масштабных вторжений вредителей за последние десятилетия. Паразитический клещ распространился по ульям медоносных пчел (Apis mellifera) по всему миру и стал причиной резкого увеличения смертности пчелиных семей, особенно в сочетании с переносимыми им вирусами. Поэтому подавляющее большинство пчеловодов предпочитают обрабатывать свои ульи от Varroa destructor, часто по несколько раз за сезон. Согласно последнему исследованию потерь пчелиных семей в Новой Зеландии, в 2023 году более 97% всех ульев были обработаны от клеща, при этом более чем в 80% случаев использовались синтетические акарициды.
Высокая вирулентность и быстрое размножение клеща требуют регулярного проведения обработок. Самки-основательницы Varroa destructor совершают свой репродуктивный цикл внутри расплодной ячейки медоносной пчелы менее чем за 12 дней. Они проникают в расплодные ячейки с личинками 5 дневного возраста непосредственно перед их запечатыванием пчелами, где питаются гемолимфой личинок, и начинают откладывать яйца примерно через 3 дня после проникновения. В течение 12 дней потомство клеща созревает и превращается во взрослых особей, сразу способных к дальнейшему размножению. Как только вылупится полностью сформировавшаяся медоносная пчела, вместе с ней появится новое поколение клещей. Такая высокая скорость воспроизводства позволяет популяциям Varroa destructor быстро расти, и, согласно моделям динамики популяций, их численность может увеличиться в 12 раз за один сезон, а в более теплом климате, где расплод присутствует постоянно, — еще больше. Чтобы предотвратить экспоненциальный рост и не допустить превышения пороговых значений экономического ущерба, пчеловоды проводят обработку несколько раз в год.
В настоящее время для борьбы с клещами используется целый ряд различных средств, часто в сочетании друг с другом, чтобы добиться максимального результата и свести к минимуму их вредное воздействие. Наиболее распространенными химическими средствами являются синтетические акарициды, такие как пиретроиды, амитаз и фосфорорганические соединения, благодаря их эффективности и простоте применения. Однако из-за их липофильной природы они легче накапливаются в улье. Такое накопление может быть опасным по разным причинам: оно приводит к повышенному воздействию и потенциальному сублетальному воздействию на пчел, а также к более высокому риску развития резистентности к акарицидам у Varroa destructor.
Все более популярной альтернативой становятся органические средства на основе эфирных масел (тимол, ментол) или органических кислот (муравьиная кислота, щавелевая кислота). Несмотря на то, что эти средства могут быть эффективными, результат их применения зачастую менее предсказуем, и даже органические продукты могут негативно влиять на расплод и здоровье пчел. Помимо химических средств, улучшить борьбу с клещами можно с помощью правильного ухода за ульем. Сетчатые донья для улавливания клещей, разделение семей, плановые перерывы в выращивании расплода или удаление трутневого расплода могут помочь снизить численность клещей в краткосрочной перспективе или замедлить их рост. Однако эффективного, простого в применении и безопасного метода лечения пока не существует.
Исследование
Ученые из School of Biological Sciences, Victoria University of Wellington (Новая Зеландия) и GreenLight Biosciences (США), испытвали новый препарат против Varroa destructor - , базирующийся на технологии РНК-интерференции (RNAi) - это новый подход к борьбе с клещами-разрушителями Варроа.
Ученые исследовали влияние Vadescana, нового препарата на основе дцРНК, на продолжительность жизни пчел и их пищевое поведение. Ульи, обработанные Вадесканой, сравнивали с ульями, обработанными обычными митицидными полосками, амитразом, и с необработанными ульями. При этом ульи с медоносными пчёлами были оснащены системой радиочастотной идентификации (RFID), а только что вылупившиеся пчёлы были помечены RFID-чипами, что позволило автоматически отслеживать перемещения пчёл-фуражиров в ульи и из ульев.
Помимо данных по эффективности нового препарата ученые обнаружили интересные факты в поведении и жизни пчел - закономерности их перемещений, воровства между пасеками и ульями, и жизненного цикла пчел.
Выяснилось, что продолжительность жизни у всех пчёл разная, при этом у необработанных пчёл она самая короткая. У пчёл из ульев, обработанных амитразом, продолжительность жизни самая большая, а у пчёл из ульев, обработанных вадесканой, — средняя. Анализ фуражировочного поведения показал, что обработка вадесканой повышает активность пчёл-фуражиров. Также было замечено, что пчёлы перелетают из улья в улей и разоряют ульи даже на отдалённых пасеках. На протяжении всего исследования количество клещей подсчитывалось с помощью поддонов в ульях и было наименьшим в группе, обработанной препаратом Vadescana. Результаты исследования показывают, что паразитирование Varroa destructor наносит вред пчелам, снижая их способность добывать пищу и сокращая продолжительность жизни. Обработка препаратами, напротив, увеличивает продолжительность жизни и активность пчел в поисках пищи.
Механизм работы нового препарата Vadescana заключается в использовании биопестицидов на основе технологии РНК-интерференции (РНКи) для борьбы с патогенами медоносных пчел, такими как вирус деформации крыла (ВДК), израильский вирус острого паралича (ИВОП) или клещ Varroa destructor.
РНК-интерференция — это естественный механизм подавления экспрессии генов у эукариот, который действует на посттранскрипционном уровне. Короткие фрагменты двухцепочечной РНК (дцРНК), полученные из генома или из экзогенных источников, таких как вирусы, связываются с большим ферментным комплексом (комплексом РНК-индуцированного сайленсинга), который вызывает специфическую деградацию комплементарных последовательностей РНК.
Препараты на основе РНК-интерференции оказались эффективными против клещей Varroa destructor при воздействии на определенные гены. Специфическая для Varroa destructor дцРНК, используемая в этом исследовании, разрабатывается компанией GreenLight Biosciences, Inc. под торговой маркой NORROA™. Действующее вещество называется вадескана (CAS# 2643947–26-4). Оно воздействует на ген кальмодулина в Varroa destructor и подавляет выработку яиц у клещей, тем самым препятствуя их размножению.
Большинство препаратов для борьбы с клещами применяются неоднократно в течение сезона выращивания расплода или в виде средств длительного действия, независимо от того, какой метод лечения выбирает пчеловод, что приводит к длительному воздействию этих веществ на пчел. Поэтому оценка их токсичности для пчел и степени сублетального воздействия является важнейшим шагом на пути к разработке безопасного метода лечения. Многие коммерчески доступные акарициды были протестированы на предмет местной и системной токсичности, средней летальной дозы и влияния на здоровье и экспрессию генов. Однако о влиянии новых методов борьбы с клещами, таких как продукты на основе дцРНК, на продолжительность жизни и поведение медоносных пчел не сообщалось.
В своем исследовании ученые использовали технологию радиочастотной идентификации (RFID) для отслеживания перемещений отдельных пчел в улье и за его пределами. RFID-чипы, прикрепленные к пчелам, не нарушают их естественное поведение и могут автоматически считываться электронными устройствами у входа в улей. Технология RFID успешно применяется для отслеживания медоносных пчел после непосредственного применения инсектицидов.
Нашей главной целью было изучить влияние обработки против клеща Varroa destructor на отдельных пчел. Пчел из ульев, обработанных вадесканой, сравнивали с пчелами из ульев, не обработанных от клещей, а также с пчелами из ульев, обработанных обычным акарицидом (полосками с амитазом). Чтобы определить общий уровень здоровья пчел и количественно оценить возможные сублетальные эффекты, ученые изучили три ключевых показателя здоровья. Они фиксировали продолжительность жизни и выживаемость пчел, их пищевое поведение, а также отслеживали динамику развития популяций клещей с помощью подсчета их количества на поддоне улья. В нашего эксперимента все средства применялись в ульях в соответствии с инструкциями производителя для достижения концентрации, сопоставимой с реальной ситуацией в полевых условиях. Этот эксперимент также позволил выявить закономерности перемещения пчел и воровства между пасеками и ульями.
Материалы и методы
Пасеки и ульи
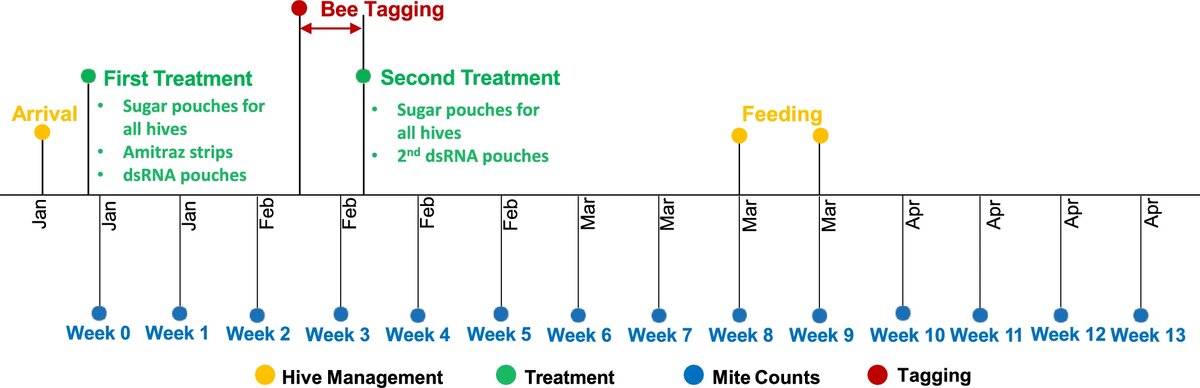
Испытания проводились в течение 17 недель с января по май 2024 года на 9 ульях, расположенных на двух разных пасеках на расстоянии 300 м друг от друга на территории Университета Виктории в Веллингтоне, Новая Зеландия (широта: −41.290, долгота: 174.767). Ульи на одной пасеке располагались на расстоянии около 2 м друг от друга, чтобы их можно было подключить к одной и той же системе радиочастотной идентификации. В начале исследования, в конце января, на территорию кампуса были привезены ульи с коммерческой пасеки (Dave’s bees, Ашерст, Новая Зеландия). На тот момент в каждом улье было по 8–10 рамок с пчелами и матка-основательница, и их не обрабатывали от клещей как минимум за 2 месяца до начала эксперимента. Все ульи были установлены на деревянное основание, чтобы под каждый улей можно было подложить поддон для контроля заражения клещами.
Все данные о погоде на этих пасеках были получены из Национальной базы климатических данных Новой Зеландии (Cliflo). Информация была получена с ближайшей метеостанции, расположенной в пределах 1 км от пасек (автоматическая метеостанция в Келбурне: широта: −41,284, долгота: 174,767), и измерялась ежедневно в 9 утра по местному времени.
Группы лечения
Ульи были разделены на три группы, в каждой из которых случайным образом назначалось одно из трех условий.
Одна группа из 3 ульев вообще не подвергалась обработке от клещей и служила отрицательным контролем.
Вторая группа из 3 ульев была обработана митицидными полосками, содержащими амитраз, в качестве контрольной группы, обработанной обычным митицидом.
Третья группа из 3 ульев была обработана вадесканой в концентрации 4 г/л. Торговая марка препарата — NORROA™. Вадескана выпускается в виде раствора на основе сахарозы с концентрацией ~60 % в герметичном пакете с перфорированными отверстиями. Этот пакет помещается в улей поверх расплодного корпуса или на нижнюю доску, чтобы пчелы могли взять раствор вадесканы и распределить его по всему расплоду и развивающихся личинок.
В начале исследования (нулевая неделя) во все ульи был помещен пакет объемом 500 мл с раствором на основе ~60% сахарозы, идентичным по составу NORROA™, но без активного ингредиента — дцРНК вадесканы. Благодаря этому приложению все ульи получали одинаковый источник питания, но только в 3 ульях были пакеты с вадесканой (рис. 1). Ульи, обработанные митицидами, получали по две полоски амитраза по 500 мг амитраза на полоску (Apivar®, Veto-pharma S.A.S., Франция) на каждый расплодный корпус в соответствии с инструкцией производителя в дополнение к раствору на основе сахарозы. Через три недели всем ульям дали второй пакет с раствором, содержащим ~60% сахарозы, или с раствором, содержащим вадескану, в соответствии с инструкциями производителя.
Пчелы с RFID- меткой
Чтобы отслеживать активность и перемещения отдельных пчел, их помечали RFID-чипами (Mini-Transponder mic3®Q1.6, microsensys GmbH, Эрфурт, Германия), каждый из которых имел уникальный идентификационный номер (UID) (рис. 2). Размеры транспондера составляли 1,6 × 1,6 × 0,4 мм3, а его вес — менее 5 мг, чтобы он не мешал пчелам вести привычный образ жизни.
На третьей неделе, когда из яиц, обработанных препаратами для борьбы с клещами, должны были вылупиться пчелы, по одной рамке запечатанного расплода из каждого улья перенесли в лабораторию и оставили на ночь при температуре 34 °C в помещении с регулируемой температурой. На следующее утро только что вылупившихся пчел собрали прямо с рамок. Пчел усыпили при температуре −20 °C на 5 минут и поместили на лед для процедуры RFID-мечения. На спинную часть груди всех пчел нанесли каплю клея на основе цианоакрилата (Shelleys®, Австралия), а сверху, между крыльями, разместили RFID-транспондер. Все метки были инициализированы, привязаны к конкретному улью и проверены на работоспособность с помощью портативного RFID-считывателя.
Для объединения идентификаторов пчел в группы и определения даты их рождения использовалось программное обеспечение iID®BEE suite (microsensys GmbH, Эрфурт, Германия) в сочетании с устройством для считывания меток. Этот процесс повторялся для каждого улья до тех пор, пока примерно 150 пчел в каждом улье не были помечены RFID-чипами. После того как пчелы полностью приходили в себя, их возвращали в ульи в течение 4 часов после мечения.
Мониторинг на основе радиочастотной идентификации (RFID)
Для непрерывного мониторинга ульев и перемещений отдельных пчел все ульи были оснащены RFID-технологией (iID®BEEscience professional Bundle v313.2D, microsensys GmbH, Эрфурт, Германия). Каждый улей был оснащен RFID-считывающим устройством (iID® Reader Block — модель с держателем для антенны на крыше), установленным на небольшой платформе у входа в улей (рис. 2). Эти считывающие устройства обнаруживают и идентифицируют все RFID-чипы, проходящие через вход. Все логи содержат уникальный идентификационный номер чипа (UID), точную временную метку и направление движения. Все считыватели на пасеке были подключены к центральному RFID-контроллеру (iID®controller) с оперативной памятью 1 ГБ и USB-накопителем на 16 ГБ (intenso®, Германия) для хранения данных.
Контроль ульев
На протяжении всего исследования ульи беспокоили как можно реже, не добавляя и не убирая ульи. После окончания медосбора все ульи дважды (на 8-й и 9-й неделях) получали по 2 л ~50-процентного раствора сахарозы в качестве пищевой добавки. Раствор сахара подавали через верхнюю кормушку, чтобы не беспокоить пчел.
Чтобы отслеживать развитие популяций Varroa destructor, не тревожа ульи, исследователи еженедельно подсчитывали количество упавших на поддон под ульем клещей. Картонные листы размером 505 мм × 405 мм, соответствующие размеру поддона для улья Langstroth, были обработаны спреем с рапсовым маслом (Woolworths®, Новая Зеландия), чтобы сделать их более липкими для клещей.
Эти липкие подложки помещали под все ульи на 24 часа, затем подсчитывали количество клещей на подложке и усредняли показатели по всем ульям в каждой группе. Хотя существуют более эффективные методы отбора проб для исследования клещей, мы использовали подложки, потому что это наименее инвазивный способ подсчета количества клещей, а мы хотели избежать отбора проб и травмирования пчел с RFID-метками в ходе эксперимента. Любой стресс или нарушение привычного образа жизни, вызванные альтернативными методами сбора клещей, могли повлиять на продолжительность их жизни и пищевое поведение, что могло исказить наши результаты.
Анализ данных
Все статистические анализы проводились с использованием R версии 4.3.1 (R Development Core Team, 2023) и RStudio (RStudio/2023.06.1, Posit Software PBC, США, 2023). Данные, собранные RFID-контроллерами, сохранялись за каждый день исследования и импортировались непосредственно в R, но из-за перебоев с электричеством два раза наборы данных загружались не полными. Считыватели сохраняли данные с интервалом в 15 секунд, чтобы учесть несколько обнаружений во время одного и того же полета пчелы и избежать длинной серии последовательных обнаружений. Были добавлены метаданные, содержащие информацию о дне рождения пчелы и улье, в котором она живет, и объединены с ежедневными записями. Ограничений по продолжительности полета не устанавливалось, и записи RFID не удалялись из логов.
Все доступные данные были разделены на две категории: данные о полете без информации о направлении, которые использовались для общего анализа активности и продолжительности жизни, и данные с информацией о направлении (запись считывателя: вылет/прилет). На основе последних был составлен список всех полетов каждой пчелы за всю ее жизнь.
Продолжительность жизни всех особей определялась как время между датой их рождения и моментом, когда они в последний раз были обнаружены RFID-считывателем. Для анализа выживаемости эти данные использовались для построения графиков выживаемости Каплана — Мейера с помощью пакетов survival и survminer (survival 3.6–4, survminer 0.4.9).
Анализ фуражировки проводился только на основе данных, содержащих информацию о направлении. В алгоритме отдельный вылет определялся как соответствующая пара записей данных: первая — «отправление», вторая — «прибытие» с одним и тем же идентификатором. На основе этих отдельных данных рассчитывалось количество и продолжительность вылетов на одну пчелу. Начало и продолжительность фуражировки были определены на основе одного и того же набора данных. Датой начала фуражировки считалось время первого вылета пчелы, а продолжительностью фуражировки — разница во времени между первым и последним вылетом. Продолжительность фуражировки в этом более широком определении включает в себя все действия медоносной пчелы вне улья, в том числе короткий подготовительный этап с ориентационными или ознакомительными полетами, а также собственно фуражировку.
Поскольку все пчелы были привязаны к своему родному улью, можно было фиксировать их перемещения в другие ульи. Если пчелы навсегда переселялись в другой улей и не возвращались в свой родной улей, их считали бродячими. Если же они возвращались в свой родной улей, их считали пчелами-воронами51. Сетевые графики для данных о грабежах и дрейфе судов были созданы с помощью пакета igraph (igraph 2.0.3.9019)52.
Для анализа динамики клещевой нагрузки в различных группах лечения была построена модель линейной регрессии с использованием функции lm из пакета lme453. В модели в качестве факторов учитывались вид лечения и количество недель, прошедших с начала исследования. Затем с помощью пакета emmeans были проведены попарные сравнения и рассчитаны предельные средние значения, необходимые для выявления существенных различий в апостериорном анализе54.
Поскольку все пчёлы были закреплены за своим родным ульем, можно было фиксировать их вылеты в другие ульи. Если пчёлы окончательно переходили в другой улей и не возвращались в родной, их относили к категории блуждающих (drifting bees). Если же они возвращались после посещения в свой родной улей, их классифицировали как воровок (robbing bees). Сетевые графики для данных о воровстве и блуждании были построены с использованием пакета igraph.
Результаты
Ульи наблюдались в общей сложности в течение 105 дней в период австралийского лета и осени — с февраля по май 2024 года. По завершении испытания все ульи оказались живыми и по‑прежнему активными.
Всего 1396 пчёл из 9 ульев были помечены RFID‑чипами. Из них 1327 пчёл хотя бы однажды были зафиксированы RFID‑считывателями.
В общей сложности 977 пчёл (69,99 %) хотя бы раз в жизни выходили на сбор корма.
Выяснилось, что вторая антенна на одном из RFID‑считывателей (улей «жёлтый 3») была неисправна — из‑за этого в записях отсутствовали данные о направлении движения. В связи с этим данный улей впоследствии был исключён из анализа фуражировочной активности. В итоговый анализ вошли только данные по пчёлам из необработанных ульев 1 и 2.
Продолжительность жизни
Время выживания (продолжительность жизни) было рассчитано для всех пчёл (n = 1327), которые хотя бы однажды были зафиксированы в своём улье.
Средняя продолжительность жизни достоверно различалась между группами с разными видами обработки. При этом наблюдалась также значительная вариабельность внутри каждой группы обработки — между отдельными ульями (см. рис. 3).
Получены следующие результаты по группам:
- Пчёлы из необработанных ульев (n = 439) имели самую короткую продолжительность жизни — в среднем 21,8 дня [95% в интервале = 20,5–23,2 дня].
- Пчёлы из ульев, обработанных вадесканой (vadescana, n = 430), занимали промежуточное положение: средняя продолжительность жизни составила 25,4 дня [95% в интервале = 24,0–26,8 дня].
- Пчёлы из ульев, обработанных амитразом (amitraz, n = 458), имели наибольшую среднюю продолжительность жизни — 28,7 дня [95% в интервале = 27,4–30,1 дня].
Различия в продолжительности жизни между группами с разными видами обработки были наиболее заметны в первые 30 дней испытания. Более низкая средняя продолжительность жизни в контрольной группе в первую очередь объяснялась более высокими начальными показателями смертности (см. рис. 3). Во всех группах обработки наблюдалась вариабельность выживаемости: отдельные пчёлы жили более чем в два раза дольше среднего показателя. Самая старая пчела в ходе испытания прожила 86 дней.
Активность
Ежедневные перемещения пчёл из всех ульев фиксировались — это позволяло убедиться, что RFID‑оборудование исправно и функционирует на протяжении всего испытания.
Кривая активности была сопоставлена с метеорологическими данными ближайшей метеостанции (расположенной в 700 м от места исследования), чтобы выявить взаимосвязь активности пчёл с внешними факторами — например, с понижением температуры воздуха или выпадением осадков.
На графике видно, что спады активности соответствуют периодам снижения температуры или выпадения осадков (см. рис. 4).
Уровни активности в целом отражают жизненный цикл рабочей пчелы, и помеченные пчёлы, как было показано, проходили через эти типичные фазы поведения. В первые 10 дней фиксировалась низкая активность — вероятно, в этот период пчёлы в основном выполняли внутриульевые задачи в роли пчёл‑кормилиц или домашних пчёл. Затем активность возрастала: пчёлы отправлялись на первые ориентировочные вылеты за пределы улья. За этим ростом перемещений следовал весьма активный период фуражировки продолжительностью 4 недели, после чего активность начинала снижаться — это свидетельствовало о гибели пчёл. Пчёлы из необработанных ульев проходили эти жизненные стадии быстрее: их активность рано достигала пика, а затем начинала снижаться. Более 98 % (n = 433) необработанных пчёл погибли спустя 7 недель. Активность в ульях, обработанных вадесканой (vadescana), фиксировалась в течение 11 недель, а последняя пчела из улья, обработанного амитразом, погибла спустя 14 недель наблюдений.
Поведение при поиске пищи
Для анализа фуражировочного поведения использовали только данные с информацией о направлении движения. При фильтрации доступных данных выяснилось, что считыватель одного из необработанных ульев (Y3) неисправен — поэтому этот улей исключили из анализа фуражировки. В общей сложности в анализ включили данные о 31 257 вылетах 977 пчёл.
Пчёлы из ульев, обработанных вадесканой, начинали фуражировку раньше всех — в возрасте 8,5 дней [95 % интервал = 8,20–8,79 дня], пчёлы, обработанные амитразом, занимали промежуточное положение с началом фуражировки в 10,4 дня [95 % интервал = 9,88–10,9 дня], а необработанные пчёлы приступали к фуражировке позже всех — в 12,6 дня [95 % интервал = 11,6–13,5 дня] (см. рис. 5).
Различия между всеми группами оказались статистически значимыми. Общая продолжительность периода фуражировки была схожей у групп, получавших обработку против клещей: ульи с вадесканой имели фуражировочный период продолжительностью 19,3 дня [95 % интервал = 18,1–20,5 дня], а ульи с полосками амитраза — 20,9 дня [95 % интервал = 19,8–22,1 дня]. Для необработанных ульев зафиксированная продолжительность фуражировочного периода оказалась самой короткой — 14,2 дня [95 % интервал = 12,7–15,6 дня], при этом время, которое необработанные пчёлы проводили в роли фуражиров, было статистически достоверно короче, чем у двух других групп. Статистически значимого различия между группами амитраза и вадесканы не выявлено (p = 0,2). В обеих обработанных группах встречались особи чьи показатели значительно отклоняются от средних значений по группе и выходят за рамки типичной закономерности, они которые фуражировали более 50 дней — то есть дольше, чем вдвое превышали среднюю продолжительность фуражировочного периода.
По интенсивности фуражировки пчёлы из ульев, обработанных вадесканой (vadescana), совершали наибольшее число вылетов — в среднем 41,1 вылета на пчелу [95 % интервал = 36,9–45,2] — и при этом имели самую короткую продолжительность вылетов: 59,7 минуты в среднем [95 % интервал = 48,5–71,0] (см. рис. 6).
Пчёлы из ульев, обработанных амитразом, занимали промежуточное положение: 30,4 вылета на пчелу [95 % интервал = 27,3–33,4] и средняя продолжительность вылета 89,0 минут [95 % интервал = 71,2–106,9].
Наименее продуктивными в рамках данного исследования оказались необработанные пчёлы из контрольных ульев: они совершали лишь 17,3 вылета на пчелу [95 % интервал = 14,1–20,5] и при этом демонстрировали самую длительную фуражировочную активность — в среднем 371,5 минуты [95 % интервал = 271,3–471,6].
Среднее количество вылетов на пчелу статистически значимо различалось между всеми группами обработки. Продолжительность вылетов оказалась достоверно больше только для необработанной контрольной группы по сравнению с обеими обработанными группами. В необработанных ульях было значительно больше особей с показателями, резко отклоняющимися от средних значений, и некоторые необработанные пчёлы демонстрировали необычное фуражировочное поведение: они совершали мало вылетов, но каждый из них был очень длительным — в частности, свыше 24 часов, что указывает на то, что такие пчёлы не возвращались в улей на ночь.
Перемещения между ульями.
Пчёлы перемещались между всеми ульями и обеими пасеками. Около 8 % (n = 113) всех помеченных пчёл посещали ульи, отличные от их родного улья. Если пчёлы навсегда покидали свой родной улей, их относили к категории «переселившиеся пчёлы» (drifting bees), тогда как пчёлы, которые возвращались в родной улей после визита в другой улей, классифицировались как «воровки» (robbing bees).
Переселившиеся пчёлы (n = 50) начинали перемещаться между ульями в более раннем возрасте — в среднем в 18,1 дня [95 % интервал = 14,4–21,7 дня], тогда как грабительское поведение (n = 63) могло начаться в любое время, чаще в более позднем возрасте — в среднем в 26,9 дня [95 % интервал = 23,7–30,1 дня]. Выявленная разница в возрасте между переселившимися и воровками оказалась статистически значимой.
На уровне ульев (при учёте более чем одной пчелы) переселение происходило только между соседними ульями на одной пасеке и в целом отмечалось на более низком уровне, чем воровство (см. рис. 7a). Такая закономерность указывает на то, что переселение может быть обычным явлением между ульями на одной пасеке: между ульями происходит обмен пчёлами (в данном случае около 3,5 %), но это скорее серия случаев, когда отдельные пчёлы случайно переселяются в соседний улей, а не скоординированное роевое явление.
Воровская активность, напротив, исходила в основном из одного улья (Red 3) и была зафиксирована в 7 из 8 других ульев — в том числе на другой пасечной площадке (см. рис. 7b). При сопоставлении данных о подобном поведении с другими наборами данных — например, о фуражировочных вылетах и продолжительности жизни пчёл — выяснилось, что некоторые вылеты для воровства происходили в один и тот же день и в одном и том же улье, а значит, были частью более скоординированных действий. В дни такого активного воровства общее число воровских и фуражировочных вылетов было выше обычного, пчёлы проявляли необычную активность, а также фиксировалось необычно большое число погибших пчёл.
Количество клещей
Для изучения влияния различных методов обработки на популяцию клещей в течение всего периода исследования еженедельно проводился подсчет клещей на доньях ульев (рис. 8). Хотя этот метод мониторинга клещей не представлял опасности для пчел с метками и практически не нарушал работу ульев, известно, что он менее точен, чем другие диагностические инструменты. На нулевой неделе, в начале исследования, во всех ульях было умеренное количество клещей — в среднем около 5 клещей на 100 пчел. Количество клещей оставалось ниже исходного уровня в среднем до 8-й недели, когда в ульях без обработки в среднем было 6,67 клеща на 100 пчел. Количество клещей во всех ульях продолжало расти, и на 10-й неделе даже в ульях, обработанных вадесканой, в среднем было 5,33 клеща на 100 пчел.
Линейная регрессионная модель, включающая основные эффекты факторов «Обработка» (F = 4,12, df = 2, p = 0,019) и «Неделя» (F = 15,83, df = 13, p < 0,001) в отношении численности Varroa destructor, показала, что оба фактора оказывают значимое влияние на количество клещей, собранных на поддонах.
Дальнейший анализ с попарными сравнениями выявил, что численность клещей в необработанных ульях была достоверно выше, чем в ульях, обработанных вадесканой (p = 0,014). В ульях, обработанных амитразом, количество клещей имело промежуточные значения — они статистически не отличались ни от необработанных ульев, ни от ульев с обработкой вадесканой (p > 0,3).
Также было установлено, что влияние фактора «Неделя» на численность клещей становится значимым начиная с 11‑й недели (p < 0,05). Такая динамика указывает на то, что обработки эффективно сдерживали уровень клещей примерно в течение 10 недель, а после их действие ослабевало.
Экспоненциальный рост численности клещей также типичен для данного сезона: осенью количество пчёл снижается, а паразитарная нагрузка со стороны клещей, напротив, резко возрастает.
Обсуждение
В данном исследовании ученые изучали влияние различных обработок против Varroa destructor на отдельных пчёл. Пчёлы из ульев, которые не обрабатывались против Varroa destructor, демонстрировали сниженную продолжительность жизни и ухудшенную фуражировочную активность. Обе обработки для борьбы с клещами — то есть полоски с вадесканой и амитразом — улучшали продолжительность жизни и здоровье пчёл по сравнению с необработанной контрольной группой.
Пчёлы, обработанные вадесканой, жили значительно дольше, чем необработанные, и тратили больше времени на выполнение задач за пределами улья — на предфуражировочной и фуражировочной стадиях. За свою жизнь они совершали больше вылетов и в среднем имели самую короткую продолжительность вылетов, тогда как необработанные пчёлы демонстрировали противоположное поведение — то есть совершали меньше вылетов, но каждый из них был более длительным. Более длительные периоды отсутствия в улье, особенно в ночное время, можно интерпретировать как признак потери ориентации и потенциального когнитивного нарушения у этих пчёл.
Поскольку ранее было показано, что пчёлы, заражённые Varroa destructor, проводят больше времени вне улья, имеют более длительные полёты и более низкую частоту возвращений, результаты, полученные для необработанной группы, можно объяснить более высоким уровнем заражения клещами. Их сокращённая продолжительность жизни также может быть связана с более низкой частотой возвращений, негативным влиянием высокого уровня заражения клещами и, возможно, вирусными инфекциями, переносимыми Varroa destructor.
Множество исследований ранее демонстрировало, что у пчёл, поражённых паразитом Varroa destructor, продолжительность жизни короче, а масса тела ниже.
Фуражировочная активность и продолжительность жизни пчёл, обработанных вадесканой, вероятно, также связаны между собой. Задачи за пределами улья, такие как ориентационные или фуражировочные полёты, как известно, связаны с различными рисками и более высокой смертностью по сравнению с работой внутри улья. Более ранний переход к фуражированию и более высокая вовлечённость в фуражировочную деятельность могут идти за счёт сокращения продолжительности жизни. Эта отрицательная корреляция может объяснить, почему пчёлы, обработанные dsRNA, совершали за свою жизнь больше вылетов, чем пчёлы, обработанные амитразом, хотя и имели более короткую среднюю продолжительность жизни. Пчёлы из обеих обработанных групп демонстрировали нормальное фуражировочное поведение: количество и продолжительность вылетов соответствовали результатам других исследований с аналогичной RFID‑установкой.
Ульи, обработанные вадесканой, постоянно демонстрировали более низкий уровень заражения клещами, что указывает на эффективность обработки. В силу механизма действия вадескана, по‑видимому, не является летальной для клещей — поэтому не ожидается увеличения падения клещей; вместо этого она снижает их репродукцию, в результате чего останавливает рост популяции и сокращает число клещей в каждой колонии.
Обработка амитразом не привела к значимому изменению падения клещей по сравнению с необработанными ульями. Хотя подсчёты клещей на поддонах менее точны, чем другие методы подсчёта, еженедельные замеры в течение длительного периода позволяют оценить динамику численности клещей и полезны для сравнения колоний в рамках эксперимента.
Еженедельные подсчёты клещей показали схожую тенденцию для всех групп обработки, что, скорее всего, связано с сезонными изменениями численности пчёл и клещей осенью. Небольшой размер семей, рост численности клещей и повторное заражение из соседних ульев могут привести к быстрому увеличению числа клещей. К концу испытания, в мае, во всех ульях численность клещей достигла порогового значения, требующего обработки.
Перемещения пчёл между ульями могут способствовать распространению клещей и росту уровня заражения. В нашем случае около 8 % помеченных пчёл переместились в соседние ульи. Мы выявили два отчётливо различающихся паттерна.
«Переселившиеся» пчёлы (drifting bees) навсегда переселялись в чужой улей, обычно в пределах одной пасеки и в более молодом возрасте — что, возможно, говорит о том, что они терялись во время ориентационных полётов.
Напротив, пчёлы‑воровки (robbing bees) посещали ульи как на другой пасеке, так и на своей, но затем возвращались в родной улей; их вылеты не имели чёткой возрастной закономерности. Воровство инициируется на уровне улья: пчёлы‑воровки привлекают других фуражиров после того, как находят подходящий целевой улей. Ульи‑воровок характеризуются изменениями в фуражировочном поведении и поведении охранных пчёл.
Во время эпизодов воровства активность фуражирования, агрессия и, следовательно, риск смертности быстро возрастают. Эти изменения также наблюдались в данных по активности и продолжительности жизни: зафиксированные эпизоды воровства соответствовали повышенной активности и смертности в улье‑грабителе.
Помимо прямых рисков — драк с пчёлами улья‑жертвы и потенциально смертельных травм — эпизоды воровства могут иметь долгосрочные последствия для здоровья и выживаемости улья. Клещи и другие патогены, такие как американский гнилец (AFB) или вирус деформации крыла (DWV), могут передаваться горизонтально в другие ульи через посещающих их пчёл. Повторное заражение ранее обработанных ульев клещами из соседних колоний может свести на нет результаты предыдущих обработок. Таким образом, перемещения пчёл между ульями могут оказывать значительное влияние на здоровье колонии и уровень заражения клещами.
Пчёлы из ульев, обработанных амитразом и вадесканой, имели среднюю продолжительность жизни 28 и 25 дней соответственно, что укладывается в обычно приводимый нормальный диапазон для медоносной пчелы — 25–35 дней.
В литературе, посвящённой конкретно продолжительности жизни пчёл с RFID‑метками, обычно сообщается о несколько более короткой продолжительности жизни — около 20 дней, даже для контрольных групп. Скорее всего, это связано с тем, что манипуляции с пчёлами и их маркировка представляют собой дополнительный стрессовый фактор, которого невозможно полностью избежать. Соответственно, даже 21 день для необработанных ульев кажется находящимся в пределах естественного диапазона.
Исследователи не обнаружили какого‑либо негативного влияния амитраза на пчёл — в отличие от других исследований, сообщающих о воздействии акарицидов, таких как флуметрин или амитраз, на здоровье и поведение пчёл. Чтобы выявить такие эффекты, пестициды или акарициды часто экспериментально вводят отдельным пчёлам напрямую — перорально либо через контакт с кутикулой. Однако, как показывают данные на уровне колонии, в полевых условиях эти вещества могут не оказывать влияния на работоспособность пчёл.
Медоносные пчёлы, обработанные вадесканой, демонстрировали достоверно более раннюю и усиленную фуражировочную активность по сравнению с пчёлами, обработанными амитразом. Как уже обсуждалось ранее, фуражировочные вылеты за пределы улья и даже ориентационные полёты, как известно, связаны с более высокой смертностью по сравнению с работой внутри улья. Более ранний переход к фуражированию может происходить за счёт сокращения продолжительности жизни. Такое фуражировочное поведение, возможно, и способствовало несколько более короткой продолжительности жизни пчёл, обработанных вадесканой, по сравнению с пчёлами, обработанными амитразом.
Обработки на основе РНКи (RNAi) представляют собой новую группу биопестицидов, которые действуют по иному механизму, чем традиционные акарициды. Поэтому их можно использовать для регулирования популяций, выработавших устойчивость к другим акарицидам, а также в качестве важного инструмента для предотвращения или замедления формирования устойчивости у популяций, ещё не обладающих резистентностью к иным акарицидам.
Комплексное управление устойчивостью предполагает, что чередование средств с разными механизмами действия может замедлить или предотвратить развитие резистентности. Смена синтетических акарицидов на акарициды на основе РНКи способна помочь пчеловодам, сталкивающимся с устойчивыми популяциями клещей в своих колониях.
Обработки с использованием дцРНК (dsRNA) успешно применялись для контроля популяций Varroa destructor в небольших масштабах. Однако данные об их воздействии в полевых условиях и на протяжении длительных периодов времени пока ограничены.
В данном исследовании обработки применялись к полноценным семьям медоносных пчёл (8–10 рамок с пчёлами) в соответствии с инструкциями производителя. Таким образом, пчёлы и клещи подвергались воздействию концентраций, сопоставимых с условиями в полевых условиях. Следовательно, полученные результаты отражают то, чего пчеловоды могут ожидать с точки зрения эффективности и влияния на фуражировочную активность в реальной практике.
Кроме того, необходимо учитывать влияние РНКи‑технологии на виды‑хозяева клещей и других насекомых, обитающих в среде обитания клещей. При воздействии вадесканы на монарховых бабочек (Danaus plexippus) и восковых молей (Galleria mellonella) негативных эффектов для этих видов выявлено не было. Предыдущие исследования также не выявили влияния вадесканы на куколок медоносных пчёл.
В соответствии с этими результатами мы не наблюдали негативных последствий для взрослых особей в отношении их фуражировочной активности или продолжительности жизни. Напротив, пчёлы, обработанные вадесканой, жили дольше, чем пчёлы из необработанных ульев, и демонстрировали более раннюю и усиленную фуражировочную активность по сравнению с пчёлами из необработанных ульев или ульев, обработанных амитразом.
Авторы:
Джоана Мерк, Маэль Анастаси, Роуз МакГрудди, Антуан Фелден и Филип Дж. Лестер - Школа биологических наук, Веллингтонский университет Виктории, Веллингтон, Новая Зеландия
Брайан Мэнли - GreenLight Biosciences, Research Triangle Park, Дарем, Северная Каролина, США
Оригинал статьи Scientific Reports https://doi.org/10.1038/s41598-026-38557-w