Трансляция - (лат. передача) - перевод нуклеотидной последовательности и-РНК в аминокислотную последовательность полипептида. Является вторым этапом реализации генетической информации и происходит непосредственно на рибосомах.
1. Подготовительный этап
- активизация аминокислот
Каждая отдельная аминокислота взаимодействует с АТФ и образует с ней активированный комплекс, который получил название аминоациладенилат. Взаимодействие происходит с помощью фермента аминоацил-т-РНК-синтетазы, специфичного для каждой аминокислоты и участвующего в следующей стадии.
- аминоацилирование т-РНК
Под действием ферментов аминоацил-т-РНК-синтетаз (кодаз) происходит присоединение аминоациладенилатов к своим т-РНК.
Фермент узнает свою т-РНК и присоединяет к акцепторной ветви молекулы определенную аминокислоту в виде аминоациладенилата. Энергия АТФ идет на образование ковалентной связи между аминокислотой и т-РНК. Аминокислота, присоединенная к акцепторной ветви кодируется кодоном, комплементарным антикодону это т-РНК.
2. Собственно трансляция
- Инициация
Малая субъединица рибосомы соединяется с и-РНК, она имеет специальный центр - аминоацильный (А-центр). Сигналом инициации служит кодон и-РНК АУГ, который считывается в А-центре. К нему у эукариот присоединяется метиониновая т-РНК, а у прокариот - формилметиониновая т-РНК.
Малая субъединица, и-РНК и т-РНК с первой аминокислотой образуют комплекс инициации.

Далее к малой субъединице рибосомы присоединяется большая и т-РНК с первой аминокислотой перемещается в соседний пептидильный центр (Р-центр), расположенный преимущественно на большой субъединице.
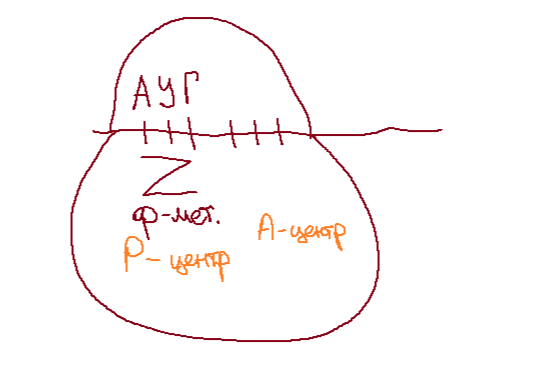
Инициация обеспечивается фактором инициации и энергией ГТФ (специфические белки).
В более редких случаях инициирующим кодоном служит валиновый кодон и-РНК ГУГ.
- Элонгация полипептида
Специальные факторы элонгации обеспечивают доставку в А-центр и закрепление в нем второй т-РНК с аминокислотой, кодируемой кодоном и-РНК, оказавшимся в А-центре.
Специальный фермент большой субъединицы рибосомы - пептидилтрансфераза осуществляет перенос первой аминокислоты из Р-центра в А-центр. Между а1 и а2 образуется пептидная связь. Теперь т-РНК в А-центре несетт дипептид, а свободная т-РНК вытесняется из рибосомы
Специальные факторы элонгации вызывают перемещение рибосомы вдоль и-РНК на один триплет, при этом система т-РНК -дипептид оказывается в соседнем Р-центре.
А-центр освобождается и факторы элонгации направляют в него и закрепляют т-РНК с третьей аминокислотой в соответствии с правилами генетического кода.
Затем пептидилтрансфераза осуществляет перенос дипептида из Р-центра в А-центр. Между а2 и а3 образуется пептидная связь и формируется трипептид. Свободная т-РНК вытесняется из рибосомы.
Происходит очередной скачок рибосомы на один триплет и система т-РНК-трипептид попадает в Р-центр.
Так рибосома последовательно идет вдоль цепи и-РНК, с каждым шагом смещаясь на один триплет, считывая с нее информацию и в соответствии с ней наращивает полипептидную цепь, начиная от ее N-конца к С-концу.
Все этапы элонгации также обеспечиваются энергией ГТФ и факторами элонгации.
- Терминация
Когда в А-центре рибосомы окажется один из нонсенс-кодонов (терминаторы: УАА, УАГ, УГА), то специальные факторы терминации узнают его и останавливают синтез полипептида. Это RF-факторы (факторы освобождения). Один из них RF1 или RF2 взаимодействуют с кодоном - терминатором, а другой фактор RF3 присоединяется к первому, а также при участии ГТФ еще и к большой субъединице рибосомы.
Связь между т-РНК и синтезируемым полипептидом гидролизуется, и полипептид освобождается от рибосомы. Происходит выход т-РНК из рибосомы и разделение субъединиц рибосомы.
Таким образом, освободившиеся субъединицы рибосом, и-РНК и т-РНК могут принимать участие в новом акте инициации трансляции.
Терминация трансляции обеспечивается факторами трансляции и энергией ГТФ.
Так как обычно клетке требуется большое количество молекул какого-либо белка, то синтез его молекул происходит на полисомах (полирибосомах), представляющих собой несколько функционирующих рибосом, объединенных одной молекулой и-РНК.
У прокариот, где ДНК не отделена ядерной мембраной от цитоплазмы и рибосом, рибосомы начинают трансляцию на цепях и-РНК, еще находящихся в стадии роста на комплексах ДНК с РНК-полимеразой. Молекулы РНК-полимеразы идут по ДНК и синтезируют цепи и-РНК, начиная от 5штрих-конца РНК в направлении к 3штрих-концу. А рибосомы присоединяются к свешивающимся с РНК-полимераз 5штрих-концевым участкам и-РНК и инициируют трансляцию, и в ходе элонгации движутся по направлению к молекуле РНК-полимеразы. Такой процесс называется сопряженной транскрипцией-трансляцией. Скорость транскрипции у бактерий - 30-45 нуклеотидов в с, скорость трансляции - 10-15 триплетов в с. Таким образом, один триплет синтезируется за то же время, за которое он считывается, и образуется пептидная связь. В рибосоме (по ее длине) может помещаться не более 30 аминокислотных остатков растущей полипептидной цепи, поэтому бОльшая часть цепи будет свешиваться в среду (цитоплазму). в ней полипептидные цепи будут скручиваться, самоорганизуясь и образуя вторичную и третичную структуры.
Гидрофобные радикалы при этом оказываются скрытыми внутри молекулы. Если же синтезируемый белок не предназначен для немедленного использования в цитоплазме, а должен первоначально транспортироваться через мембрану, то его цепь не сворачивается. В таком случае рибосомы или сами сидят на мембране и полипептид поступает через мембрану (Шероховатая ЭПС), или синтезируемая полипептидная цепь по мере ее выхода из рибосомы взаимодействует со специальными белками - молекулярными шаперонами.