Транскрипция - (лат. переписывание) - процесс перевода нуклеотидной последовательности ДНК в нуклеотидную последовательность РНК, то есть синтез РНК на матрице ДНК. Это первая стадия реализации генетической информации, в основе транскрипции лежит структурный принцип комплементарного спаривания оснований. Транскрипция осуществляется ферментами - ДНК-зависимыми РНК-полимеразами, синтезирующими РНК на ДНК матрице из рибонуклеозидтрифосфатов.
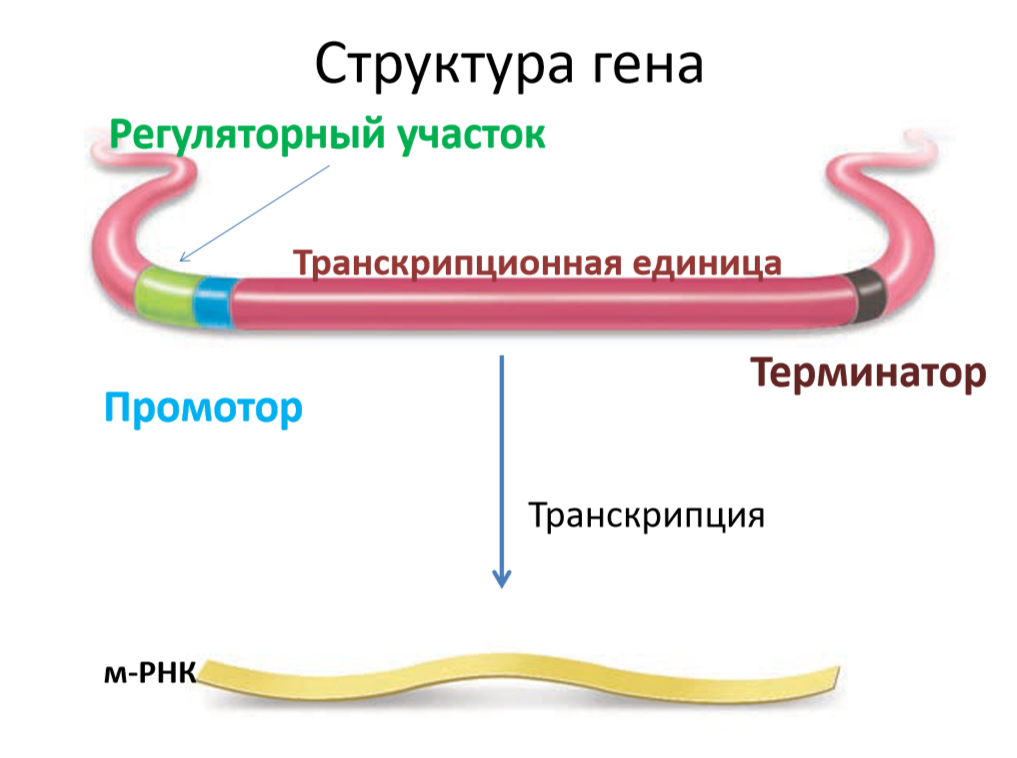
Синтез молекул РНК начинается в определенных участках ДНК - промоторах, а завершается в участках ДНК - терминаторах. Участок ДНК, ограниченный промотором и терминатором, представляет собой единицу транскрипции - транскриптон. В пределах каждого транскриптона копируется только одна из двух нитей ДНК, которая называется значащей (матричной, смысловой). Разбивка молекулы ДНК на множество транскриптонов обеспечивает возможность независимого считывания разных генов, их индивидуальные включения и выключения.
У эукариот в состав транскриптона входит один ген. Транскриптоны прокариот обычно содержат по нескольку функционально связанных друг с другом генов и называются оперонами. Структурные гены оперона как правило обуславливают синтез ферментов одной цепи превращения. Существуют опероны, содержащие гены, но не кодирующие белков - гены т-РНК, гены р-РНК. Есть также смешанные опероны, включающие гены т-РНК и белков.
Характеристика РНК-полимераз
Лучше изучены РНК-полимеразы бактерий. Большой вклад в расшифровку структуры РНК-полимеразы кишечной палочки внесла группа российского ученого Хесина. У бактерий молекула РНК-полимеразы состоит из четырех субъединиц: двух идентичных субъединиц - альфа, и двух неидентичных - бетта и бетта штрих. В бетта и бетта штрих -субъединицах находится по одному атосу цинка, входящих в каталитические центры. Эти центры обеспечивают контакт РНК-полимеразы с ДНК. Функция альфа-субъединиц пока не ясна. Такой тетрамер 2альфабеттабетташтрих РНК-полимеразы называют минимальным ферментом (кор-ферментом). Минимальная РНК-полимераза способна осуществлять транскрипцию, но она не может узнавать промотор и присоединяться к нему. Для присоединения РНК-полимеразы к промотору необходимо присоединение к минимальному ферменту еще одной субъединицы Сигма-фактора. У бактерий одна и та же РНК-полимераза осуществляет транскрипцию всех видов РНК.
В клетках эукариот обнаружено три формы РНК-полимеразы. РНК-полимераза-I (А) осуществляет синтез высокомолекулярной р-РНК, РНК-полимераза-II (В) - синтез и-РНК, РНК-полимераза-III (С) - синтез т-РНК и низкомолекулярных р-РНК.
Каждая форма РНК-полимеразы состоит из двух больших субъединиц и от 5 до 13 малых. Некоторые малые субъединицы являются общими для всех форм РНК-полимераз. Большие субъединицы выполняют функции, аналогичные бетта и бетта штрих- субъединицам бактериальных РНК-полимераз, а функции малых субъединиц пока не изучены.
В самых больших субъединицах эукариотических РНК-полимераз обнаружено несколько участков, сходных по своему аминокислотному составу с бетта штрих-субъединицей. В следующих по размерам субъединицах эукариотических РНК-полимераз есть аминокислотные последовательности, сходные с бетта-субъединицей бактериальных РНК-полимераз. Эти данные позволяют предположить, что на заре эволюции эукариот, у них имелась одна форма РНК-полимеразы, а разные формы возникли за счет умножения предковых геной и впоследствии их расхождения в результате мутации.
Особые РНК-полимеразы обеспечивают транскрипцию в хлоропластах и митохондриях. Хлоропластная ДНК содержит гены, гомологичные генам, кодирующим альфа, бетта и бетта штрих - субъединицы кишечной палочки.
Сходны также промоторы бактерий и хлоропластов. Эти факты говорят о том, что РНК-полимераза хлоропластов должна быть сходной с РНК-полимеразой бактерий. По-видимому, бактерия состоит их одной субъединицы. Кодирующий ее ген находится в ядре. У архебактерий не обнаружено множество форм РНК-полимераз, но по своему субъединичному составу их РНК-полимеразы похожи на РНК-полимеразы эукариот. Они содержат от 8 до 10 субъединиц, а большие субъединицы их гомологичны соответствующим субъединицам РНК-полимераз эукариот.
Основные этапы транскрипции.
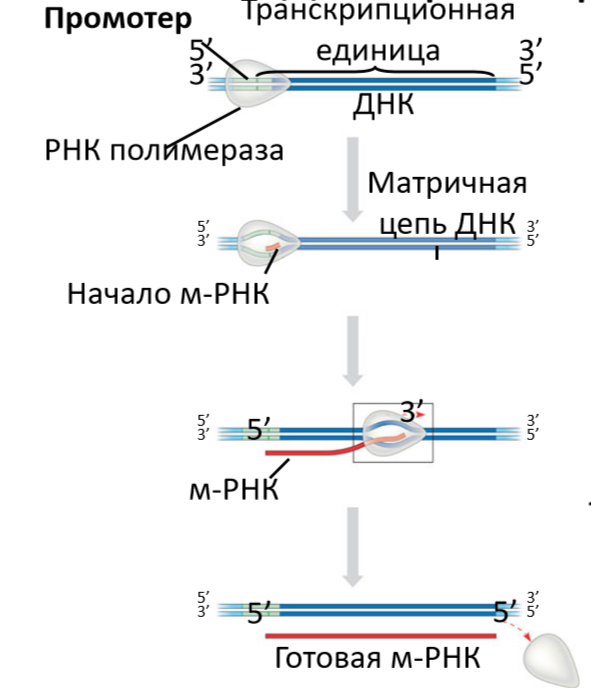
Связывание РНК-полимеразы с промотором.
Механизм поиска промотора изучен еще недостаточно. Считается, что молекула РНК-полимеразы присоединяется к случайному участку двойной спирали ДНК, некоторое время перемещается по ней, время от времени отсоединяясь и вновь присоединяясь к ней, пока не окажется на промоторе. На промоторе РНК-полимераза образует с ним первоначально закрытый комплекс, в котором ДНК еще двухспиральна. В нем РНК-полимераза не может синтезировать РНК, комплекс нестабилен и легко диссоциирует при повышении ионной силы. Закрытый комплекс может превращаться в открытый, в котором РНК-полимераза расплетает примерно один виток двойной спирали ДНК в районе стартовой точки: нуклеотида, с которого начинается комплементарное копирование матрицы. В таком комплексе связь РНК-полимеразы с промотором более прочна, чем в закрытом.
Инициация.
Заключается в образовании нескольких первых звеньев цепи РНК. Первый нуклеотид в составе цепи РНК сохраняет свою трифосфатную группировку, а последующие присоединяются к 3штрих-ОН-группе предыдущего нуклеотида с освобождением пирофосфата.
Элонгация.
Происходит постепенное наращивание цепи РНК в направлении от ее 5штрих-конца к 3штрих-концу.
Расплетенными бывают примерно 18 нуклеотидных пар. Примерно 12 нуклеотидов матричной цепи ДНК образуют гибридную спираль с растущим концом РНК. По мере движения РНК-полимеразы по матрице впереди нее происходит расплетание двойной спирали ДНК, а позади - ее восстановление. Одновременно высвобождается очередное звено растущей цепи РНК из комплекса с матрицей и РНК-полимеразы. Вероятнее всего, что для предотвращения вращения РНК-полимеразы и ДНК, движущуюся РНК-полимеразу сопровождают ферменты - топоизомеразы.
Терминация.
Завершается синтез РНК на терминаторах. У прокариот для этого к минимальному ферменту РНК-полимеразы должен присоединиться белковый фактор Ро-фактор. При этом от ДНК отделается синтезированная РНК и РНК-полимераза.
Процессинг.
Так как в ходе процесса образуются еще незрелые РНК (РНК-предшественника), то в дальнейшем происходит их дозревание - процессинг. В самых простых случаях он заключается в удалении избыточных нуклеотидных последовательностей с обеих концов молекул РНК. При процессинге т-РНК и р-РНК происходит метилирование ферментами - метилазами их азотистых оснований с образованием минорных оснований. В ходе процессинга и-РНК к ней присоединяются КЭП-фрагмент (к 5штрих-концу) и поли-А-фрагмент (к 3штрих-концу).
Такое присоединение приводит к стабилизации и-РНК, увеличивает продолжительность ее жизни и обеспечивает транспорт в цитоплазме. В ходе процессинга была открыта способность некоторых видов РНК выполнять ферментативную функцию. Например, РНК-овая часть фермента рибонуклеазы-Р, катализирующего процессинг 5штрих-концов предшественников т-РНК, при ее отделении от белковой части фермента катализирует эту реакцию. Она получила название рибозин (белковая часть).
В клетках эукариот происходит особый тип процессинга - сплайсинг, которому обязательно подвергаются все молекулы и-РНК и только некоторые (очень немногие) т-РНК и р-РНК. В молекулах такого типа между собой чередуются кодирующие зоны (экзоны) и некодирующие (интроны). Это происходит потому что ДНК эукариот в своей структурной части содержит соответствующие участки. Первоначально при транскрипции образуются незрелые и-РНК или про-и-РНК (пре-и-РНК). Молекулы незрелых и-РНК содержат оба участка (экзоны и интроны). В ходе сплайсинга происходит вырезание и удаление ферментами - эндонуклеазами интронов и сшивание лигазами разрозненных экзонов в единую цепь и-РНК.
В некоторых случаях возможны различные варианты компановки экзонов в цепь и-РНК. Такое явление носит название - альтернативный сплайсинг. Он приводит к образованию нескольких различных и-РНК, кодирующих различные белки.
Иногда в сплайсинге участвует не одна молекула РНК, в этом случае происходит образование ковалентных связей между фрагментами РНК, синтезированными на разных генах. Эти гены могут быть локализованы даже в разных хромосомах. Такое явление получило название транс-сплайсинг. Существование транс-сплайсинга и альтернативного сплайсинга сильно расширяет возможности образования достаточно разнообразной популяции клеточных РНК, кодируемых ограниченным числом генов.
Синтезированные молекулы РНК образуют комплекс с белками и транспортируются через ядерные поры в цитоплазму. Предполагают, что процессинг РНК с образованием зрелых молекул продолжается и в ходе их транспорта в составе рибонуклеопротеинных частиц через ядерные поры.