Репарация (reparatio - восстановление) - свойственный всем живым организмам процесс восстановления нативной (природной) структуры ДНК, поврежденной при нормальном биосинтезе ДНК в клетке, а также при воздействии физических и химических агентов.
Предполагают, что у первых живых организмов функцию носителя генетической информации выполняла РНК, но, видимо, уже на ранних стадиях эволюции ее заменила в этом качестве ДНК. Такому событию могла способствовать бОльшая химическая устойчивость ДНК, связанная с заменой рибозы на дезоксирибозу и двухцепочечное строение, «скрывающее» целый ряд реакционноспособных группировок. Но несмотря на это, ДНК постоянно подвергается химическим изменениям как спонтанным, так и индуцированным мутагенами и даже клеточными метаболитами. Причиной повреждения ДНК могут быть радиация, ионизирующее и ультрафиолетовое облучение. Большинство возникающих в ДНК изменений недопустимы, так как они либо приводят к вредным мутациям, либо блокируют репликацию ДНК и ведут к гибели клеток. Поэтому все клетки имеют специальные системы исправления повреждений ДНК, то есть системы репарации ДНК. Причины репарации ДНК у всех механизмов сходны, поэтому рассмотрим их на примере кишечной палочки, где они сравнительно хорошо изучены.
Нарушения, возникающие в ДНК.
Сравнительно часто и спонтанно в ДНК происходит апуринизация и дезаминирование азотистых оснований. Каждая клетка человека ежедневно теряет около 5 тыс. пуринов. Результатом апуринизации является AP-сайт. Это дезоксирибонуклеотид, лишенный основание. При дезаминировании цитозин превращается в урацил, аденин - в гипоксантин, урацил - в ксантин.
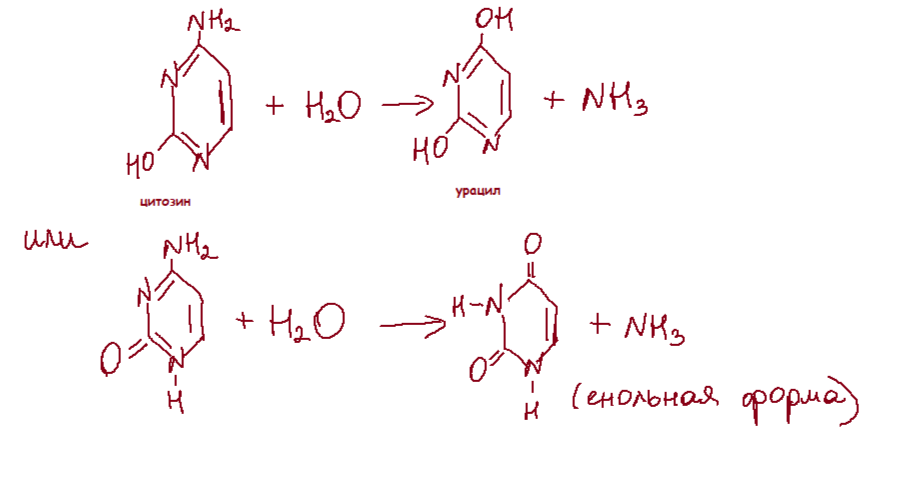
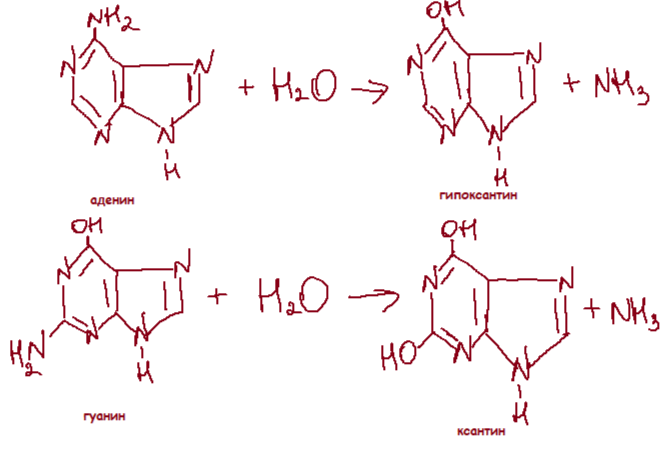
В ДНК каждой клетки человека происходит за сутки около 100 таких событий. Чаще всего дезаминируется цитозин. При дезаминировании любых оснований возникают нехарактерные для ДНК основания. Это позволяет репаративной системе клетки узнавать продукт дезаминирования и удалять его. Можно считать, что именно поэтому ДНК, в отличие от РНК, присутствует тимин, а не урацил: урацил не отличим от продукта спонтанного дезаминирования цитозина.
Многие канцерогены алкилируют основания ДНК, чаще всего метилируют. Наиболее частые продукты реакций метилирования:
О - метилгуанин, 7 - метилгуанин, 3-метиладенин
Иногда может происходить размыкание пуринового кольца гуанина. Продукт такой реакции размыкания - формамидопиримидин, затрудняет репликацию.
Главным нарушением, возникающим под действием ультрафиолета, является насыщение двойных связей оснований и образование пиримидиновых димеров из двух соседних пиримидинов цепи ДНК. Чаще всего димеров тимина.
Основные типы репарации.
1) Прямая реактивация.
Клетка устраняет ряд повреждений путем прямой реактивации. У бактерий существует специальный фермент метилтрансфераза, который приносит метильную или этильную группу с алкилированного соединения на один из собственных цистеиновых остатков.
Известна репарация повреждений, вызванных в ДНК у/ф светом, при помощи видимого света, получившая название фотореактивация (световая репарация). Ферментом служит дезоксирибопиримидинфотолиаза, способная разрезать димеры тимина до мономеров. В темноте этот фермент образует комплекс с димерами пиримидиновых оснований. На свету этот комплекс распадается и при этом происходит монодимеризация димеров. Фотореактивация широко распространена у всех групп организмов.
2) Эксцизионная репарация.
Если невозможна прямая реактивация, в клетке работают механизмы эксцизионной репарации, удаляющие из ДНК нарушенные участки. Практически на каждое аномальное основание, которое может возникнуть в ДНК существует свой специфический фермент, который называется ДНК-N-гликозилаза, узнающий в ДНК определенное аномальное основание и вырезающий с разрывом N-гликозидной связи. После его действия в ДНК остается АР-сайт. Репарация такого повреждения может идти двумя путями:
- обнаружен в клетках фермент ДНК-инсертаза, способный непосредственно пришивать к дезоксирибозе основания в соответствии с комплементарной цепью ДНК. Пока обнаружены только пуриновые инсертазы, видимо потому что апуринизация происходит намного чаще, чем потеря пиримидинов;
- связан с работой ферментов - АР-эндонуклеаз, разрывающих сахарофосфатный остов ДНК (один тип ферментов рвет фосфодиэфирную связь с 3штрих-конца от АР-сайта, другой - с 5штрих-конца). Разорванная цепь затем подвергается действию фермента - экзонуклеазы, удаляющей поврежденные участки. Затем фермент ДНК-полимераза восстанавливает поврежденный участок, а ДНК-лигаза сшивает остающийся разрыв. Общими этапами данного типа репарации являются: узнавание поврежденного участка ДНК, нарезание одной цепи ДНК вблизи повреждения (инцизия), удаление поврежденного участка ДНК (эксцизия), ресинтез ДНК, восстановление непрерывности в цепи ДНК за счет образования ковалентной связи.
При репарации повреждений ДНК, вызванных у/ф светом, поврежденные участки узнает фермент у/ф-эндонуклеаза. Вырезание осуществляет у/ф-экзонуклеаза, которая вырезает и удаляет пуриновые димеры в составе короткого олигонуклеотида. Деградация (распад) вырезанного участка ДНК - АТФ-зависимая ДНК-лигаза. Затем происходит ресинтез поврежденного участка ДНК ДНК-полимеразой и восстановление непрерывности репарируемой цепи ДНК-лигазой. Этот тип репарации повреждений ДНК, вызванных у/ф, называют темновой репарацией.
Другие типы репарации.
Если повреждений в клетке много и инцизионная репарация не справляется с устранением всех повреждений включается пострепликативная репарация. В этом случае воспроизведение в ходе репликации поврежденных молекул приводит к появлению дочерних молекул с однонитевыми пробелами, а нативная (природная) структура восстанавливается с помощью рекомбинации. Рекомбинационная пострепликативная репарация - это быстрый способ восстановления нативной структуры у дочерних молекул ДНК.
Существует также медленная пострепликативная репарация, происходящая за несколько часов. Для нее нужна система ферментов, отсутствующих в необлученных клетках и индуцируемых облучением. Этот механизм называется SOS-репарацией. Характерная его черта - неточность восстановления первичной структуры, в связи с чем он называется репарацией, склонной к ошибкам.
Нарушения репаративных процессов обнаружены у людей, пораженных наследственным заболеванием - пигментной ксеродермой, которая выражается повышенной чувствительностью к солнечному свету, приводящий к раку кожи. Известно три типа болезни: XP-I, XP-II, XPvar. У больных XP-I отсутствует у/ф-эндонуклеаза, при XP-II нет ДНК-полимеразы-I, а при XPvar наблюдается дефект пострепликативной репарации.