Проводя историко-генетические изыскания, мы подобно картографам сталкиваемся с проблемой выбора системы отсчета. Положение точки A относительно центра O можно описать множеством способов: в классической прямоугольной системе, в повернутой на 45 градусов или же в неортогональной. Каждый подход даст уникальные координаты.
Аналогичным образом, различные методики генетического анализа и выбор референтных популяций создают distinct «системы координат», прямое сравнение между которыми без учета их базовых принципов может приводить к методологическим коллизиям.
Предлагаю, отталкиваясь от этой метафоры, рассмотреть ключевые процессы в заселении Европы после Последнего ледникового максимума (ПЛМ), который служит отправной точкой для новой главы в истории континента.
Постгляциальное заселение: две прародины
Во время ПЛМ обширные пространства Центральной и Западной Европы представляли собой безжизненную пустыню, сдавленную с севера Скандинавским ледниковым щитом, а с юга и востока — альпийскими и карпатскими ледниками. Жизнь теплилась в нескольких рефугиумах, главным образом на атлантическом побережье и к северу от Пиренеев. Именно оттуда начали свою экспансию коренные обитатели Европы, пережившие оледенение, — так называемые западные охотники-собиратели (WHG – Western Hunter-Gatherers). Генетический портрет этой популяции определяли Y-хромосомная гаплогруппа I2 и митохондриальные гаплогруппы U5, U4 и U2.
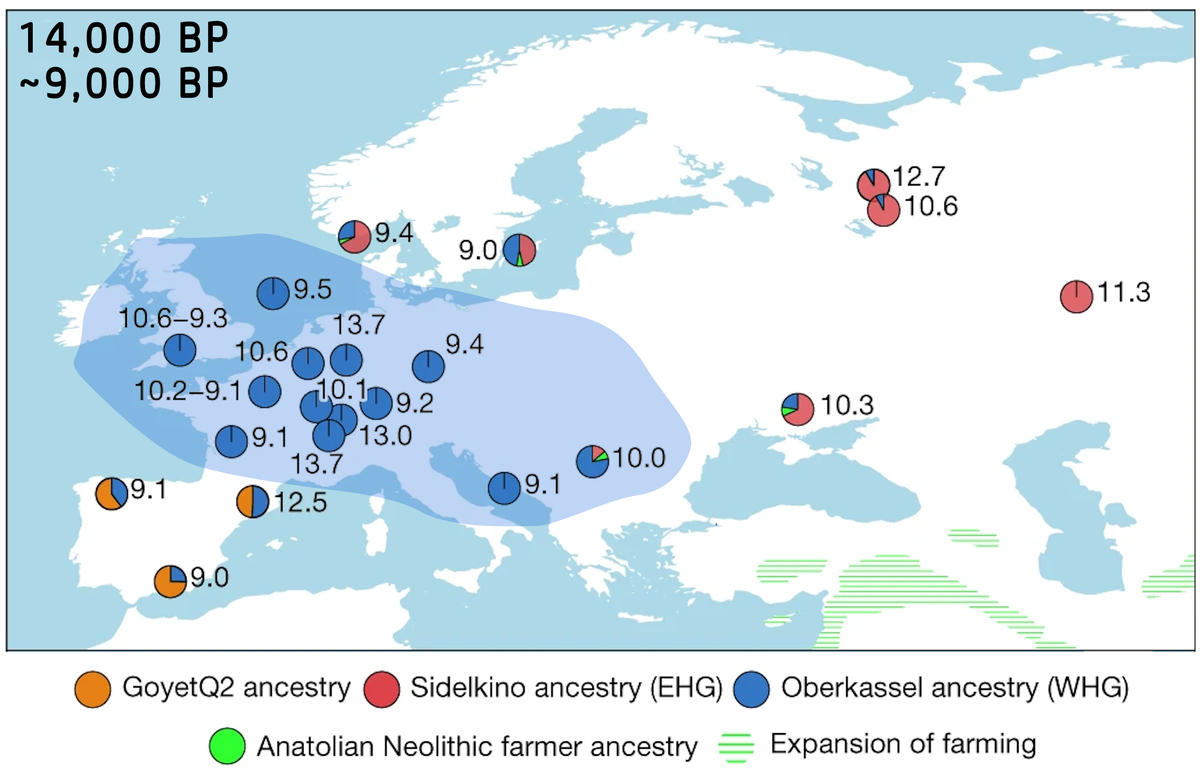
Постепенно потомки WHG заселили большую часть Западной и Центральной Европы, достигнув восточной оконечности Балтийского бассейна и тех районов Скандинавии, что освободились ото льда.
Иная картина наблюдалась в Восточной Европе. Хотя Скандинавский ледник доходил до Валдая, образуя гигантские подпрудные озера, территории к югу от современной Московской области и вплоть до Черного моря, включая Приуралье, оставались пригодными для обитания. Этот регион не только сохранил автохтонное население, но и испытывал миграционные импульсы: с запада проникали группы из Центральной Европы, а с Балкан — популяции, генетически близкие к населению Анатолии.
Затем в Восточную Европу с востока мигрировали популяции сибирского происхождения, которые со временем стали доминирующими. Так сформировался генетический кластер восточных охотников-собирателей (EHG – Eastern Hunter-Gatherers). Эти «сибиряки» были носителями североевразийского (ANE – Ancient North Eurasian) компонента, для которого характерны Y-гаплогруппы R (преимущественно R1a и R1b) и митохондриальные гаплогруппы U4, U5 и U2. Сходство мтДНК WHG и EHG является важным маркером, подтверждающим глубокие общие корни и миграционные волны из Южной Сибири в более ранние эпохи.
EHG заселили и север Восточной Европы, где в регионе Балтики и Скандинавии вступили в контакт с WHG, образовав смешанную группу — скандинавских охотников-собирателей (SHG – Scandinavian Hunter-Gatherers). При этом стоит подчеркнуть региональные различия: южные EHG активно метисировались с населением южной части Восточной Европы, в то время как северные группы сохраняли относительную генетическую обособленность вплоть до контактов в Скандинавии.
Новые генетические импульсы: земледельцы и кавказцы
Около 9 тысяч лет назад через Балканы в Европу началась массовая миграция носителей неолитических технологий — анатолийских неолитических земледельцев (ANF), позднее переклассифицированных в ранних европейских фермеров (EEF). Их генетический профиль определяли Y-гаплогруппа G2a и комплекс митохондриальных гаплогрупп: N1a, K1a, J, T, H, V, W, X и пр.
Следующей мощной волной стали мигранты с Кавказа — кавказские охотники-собиратели (CHG – Caucasus Hunter-Gatherers). Для них были характерны Y-гаплогруппы J1 и J2, а также мтДНК: K3, N1, U8, U и H13. Примечательно, что многие мтДНК-линии CHG в Европе не прижились, что указывает на преимущественно мужской характер этой миграции.

Таким образом, к началу IV тысячелетия до н.э. генетический ландшафт Европы (за исключением ее южной периферии) представлял собой мозаику из четырех основных компонентов: автохтонных WHG, восточных EHG, ближневосточных EEF и кавказских CHG, причем EHG демонстрировали внутреннюю дифференциацию на степной (южный) и лесной (северный) варианты.
Смешение и формирование новых общностей
Изначально эти генетические общности сосуществовали с минимальным смешением, однако со временем процессы метисации интенсифицировались. В Западной и Центральной Европе сформировались популяции на основе смешения WHG и EEF, ярким примером которых является культура воронковидных кубков (КВК). На севере, в Балтийском регионе, шло смешение WHG и EHG.
Наиболее динамичные процессы развернулись в степной зоне Восточной Европы, где в результате экспансии CHG начала формироваться новая степная общность — носители ямной культуры.
Аутосомный (общий геномный) состав ямников сложился как результат смешения:
· ~60% EHG (восточноевропейские охотники-собиратели)
· ~40% CHG (кавказские охотники-собиратели)
Однако мужской генофонд ямной культуры был поразительно однороден и характеризовался абсолютным доминированием Y-гаплогруппы R1b, в особенности субклада R1b1a1a (R1b-M269) и его производных, таких как R1b-Z2103. Митохондриальный же профиль в значительной степени унаследовал линии эпохи охотников-собирателей (U5, U4, U2), обогатившись пришлыми гаплогруппами от EEF: T2, T1, K, J1, W, H и другими.
Данная модель объясняет генетическое сходство между представителями ямной и шнуровой керамики культур, поскольку обе унаследовали значительную долю ANE-компонента и сходный набор сибирских мтДНК-линий.
Но остается открытым ключевой вопрос о природе кавказского компонента (CHG). Было ли его привнесение в степи исключительно односторонним процессом с Кавказа? Или же сама популяция CHG уже несла в себе существенную долю ANE, что может указывать на более глубокую общность и двусторонние миграционные потоки между Кавказом и степью?
Ответ, возможно, кроется в изначальной метафоре. Мы пытаемся сравнить данные, полученные в разных «системах координат». Без их точной калибровки и учета всех историко-демографических нюансов мы рискуем получить искаженную картину великого прошлого человечества.