Геном человека содержит 25 генов, кодирующих селенопротеины. В активном центре молекул этих белков содержится селеноцистеин; они обладают широким спектром функций: от антиоксидантной и противовоспалительной до участия в выработке активных форм гормонов щитовидной железы [Kohrle et al., 2005; Labunskyy et al., 2014].
В щитовидной железе концентрация селена максимальна по сравнению с другими органами. Тироциты экспрессируют селенопротеины - изоферменты дейодиназы (первого и второго типов, DIO1 и DIO2), глутатионпероксидазы (GPX1, GPX3 и GPX4), тиоредоксинредуктазы (TXNRD1, TXNRD2 и TXNRD3), селенопротеин 15 (SELENOF), селенопротеины P (SELENOP), M (SELENOM) и S (SELENOS) [Schomburg, 2011]. Семь селенопротеинов, включая SELENOS и SELENOK, регулируют процессы внутри эндоплазматического ретикулума. DIO1, DIO2, GPX1, GPX4, MSRB1 (метионин сульфоксид редуктаза), SEPHS2 (селенофосфат синтетаза) и TXNRD1 (рецептор тиреотропного гормона 1) представляют собой внутриклеточные селеноферменты, тогда как TXNRD2 (рецептор тиреотропного гормона 2) и GPX4 локализованы в митохондриях. Белок 2, связывающий вставку селеноцистеина (SECIS) (SECISBP2 или SBP2) – фактор, ограничивающий скорость и контролирующий трансляцию селенопротеина (воспалительные цитокины подавляют эти процессы) [Schmutzler et al., 2007].
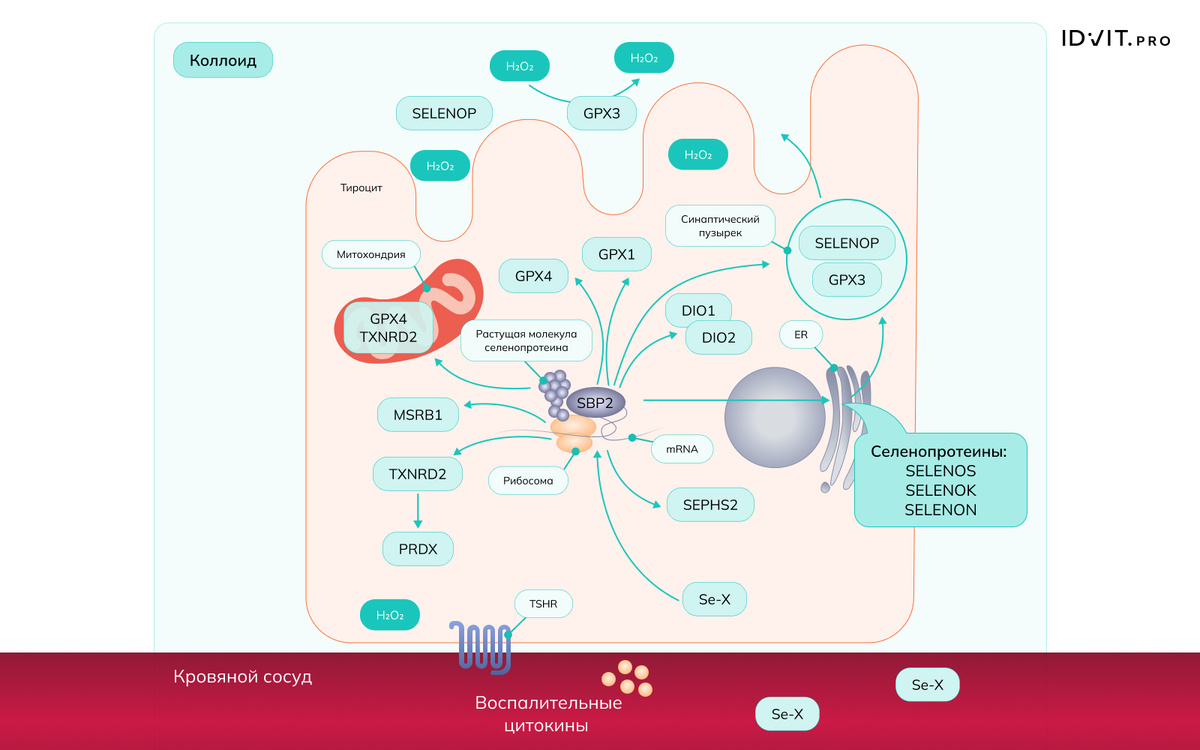
GPX3 секретируется на апикальной стороне мембраны тироцитов, где этот фермент расщепляет избыток H2O2, превращая перекись в воду. H2O2 используется тироидной пероксидазой (TPO) для йодирования тирозильных остатков тиреоглобулина и для связывания йодтирозина [Schomburg, Kohrle, 2008]. GPX1 защищает внутриклеточный компартмент от избыточного H2O2, который может диффундировать в тироциты, тогда как GPX4 может расщеплять окисленные липиды как в митохондриях, так и во всех клеточных мембранах. Низкая экспрессия и генетическая вариабельность GPX3 связаны с дифференцированным раком щитовидной железы [Lin et al., 2009]. SELENOS — селенопротеин, локализованный в ЭПР, контролирует развитие воспалительной реакции и препятствует транскрипции генов провоспалительных цитокинов, которые задействованы в патогенезе аутоиммунного тиреоидита [Santos et al., 2014]. Гомозиготность по аллелю Ala (полиморфизма DIO2 rs225014 – Thr92Ala) ассоциирована со стрессом эндоплазматического ретикулума и гипотиреозом.
Индивидуальные наследственные особенности структуры селенопротеинов влияют на предрасположенность к определенным заболеваниям (аутоиммунным и онкологическим заболеваниям щитовидной железы, кишечника и других органов, сердечно-сосудистым, инфекционным заболеваниям, нарушениям миелинизации нервных волокон и др. [Ferreira et al., 2024]), выбор терапевтического подхода к лечению и на потребности в дополнительном количестве поступающего в организм селена [Winther et al., 2020].
Ferreira, R. R., Carvalho, R. V., Coelho, L. L., Gonzaga, B. M. S., Bonecini-Almeida, M. D. G., Garzoni, L. R., & Araujo-Jorge, T. C. (2024). Current Understanding of Human Polymorphism in Selenoprotein Genes: A Review of Its Significance as a Risk Biomarker. International journal of molecular sciences, 25(3), 1402.
Köhrle, J., Jakob, F., Contempré, B., & Dumont, J. E. (2005). Selenium, the thyroid, and the endocrine system. Endocrine reviews, 26(7), 944–984.
Labunskyy, V. M., Hatfield, D. L., & Gladyshev, V. N. (2014). Selenoproteins: molecular pathways and physiological roles. Physiological reviews, 94(3), 739–777.
Lin, J. C., Kuo, W. R., Chiang, F. Y., Hsiao, P. J., Lee, K. W., Wu, C. W., & Juo, S. H. (2009). Glutathione peroxidase 3 gene polymorphisms and risk of differentiated thyroid cancer. Surgery, 145(5), 508–513.
Santos, L. R., Durães, C., Mendes, A., Prazeres, H., Alvelos, M. I., Moreira, C. S., Canedo, P., Esteves, C., Neves, C., Carvalho, D., Sobrinho-Simões, M., & Soares, P. (2014). A polymorphism in the promoter region of the selenoprotein S gene (SEPS1) contributes to Hashimoto's thyroiditis susceptibility. The Journal of clinical endocrinology and metabolism, 99(4), E719–E723.
Schmutzler, C., Mentrup, B., Schomburg, L., Hoang-Vu, C., Herzog, V., & Köhrle, J. (2007). Selenoproteins of the thyroid gland: expression, localization and possible function of glutathione peroxidase 3. Biological chemistry, 388(10), 1053–1059.
Schomburg L. (2011). Selenium, selenoproteins and the thyroid gland: interactions in health and disease. Nature reviews. Endocrinology, 8(3), 160–171.
Schomburg, L., & Köhrle, J. (2008). On the importance of selenium and iodine metabolism for thyroid hormone biosynthesis and human health. Molecular nutrition & food research, 52(11), 1235–1246.
Winther, K. H., Rayman, M. P., Bonnema, S. J., & Hegedüs, L. (2020). Selenium in thyroid disorders - essential knowledge for clinicians. Nature reviews. Endocrinology, 16(3), 165–176.