Хиральность, или "зеркальность" (от др.-греч. χείρ — рука), — фундаментальное свойство, проявляющееся на всех уровнях, от молекул до целых организмов. Аминокислоты, из которых строятся наши белки, в подавляющем большинстве "левые" (L-изомеры), а сахара — "правые" (D-изомеры). У брюхоногих моллюсков (Gastropoda) хиральность проявляется наиболее очевидно: в направлении закрученности раковины.

Большинство видов (около 90%) являются декстральными (правозакрученными, от лат. dexter — правый). Если держать такую раковину устьем к себе, оно будет расположено справа от центральной оси. Гораздо реже встречаются виды, у которых все особи синистральные (левозакрученные, от лат. sinister — левый).
Наследование этого признака — классический пример неменделевской генетики, который ставил в тупик ранних исследователей. Он демонстрирует явление, известное как материнский эффект. Классическим объектом для изучения этого феномена стал пресноводный прудовик Lymnaea peregra (или родственный ему Lymnaea stagnalis). Этот моллюск оказался идеальной моделью, поскольку он является гермафродитом и, что важно, способен как к перекрестному оплодотворению, так и к самооплодотворению.
Загадочные скрещивания: Куда пропало расщепление?
Чтобы понять этот механизм, крайне важно рассмотреть два реципрокных скрещивания. Обозначим доминантный аллель правозакрученности D, а рецессивный аллель левозакрученности — d.
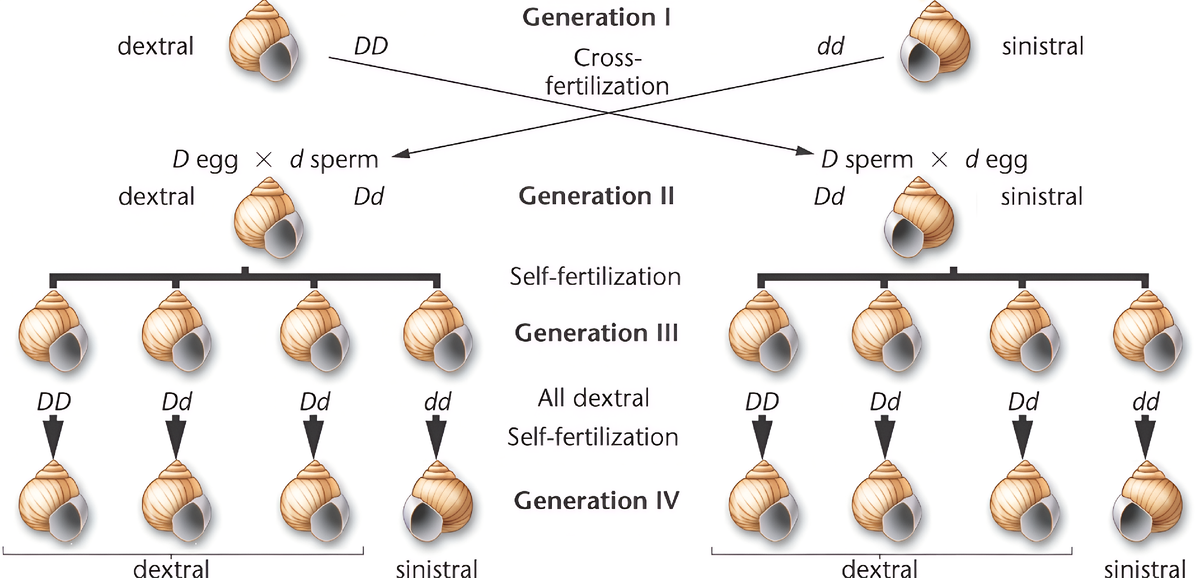
Скрещивание 1: Правозакрученная мать
P (Родители):
- Мать (♀): DD (фенотип: правозакрученная)
- Отец (♂): dd (фенотип: левозакрученный)
F1 (Первое поколение):
- Генотип: Все особи Dd.
- Фенотип: Все 100% особей правозакрученные.
Объяснение: Это поколение развивается из яйцеклеток, произведенных матерью DD. Ее генотип определяет фенотип F1.
F1 x F1 (Самооплодотворение):
Позволяем особям F1 (Dd) размножаться.
F2 (Второе поколение):
- Генотипы: 1 DD : 2 Dd : 1 dd (классическое менделевское расщепление по генотипу).
- Реальный фенотип F2: Все 100% особей — правозакрученные.
Объяснение: Все поколение F2 развивается из яйцеклеток, произведенных матерью F1 (генотип Dd). Поскольку у нее есть доминантный аллель D, она производит "правозакрученную" мРНК и белки, которые определяют фенотип всего ее потомства, включая тех особей, что получили генотип dd.
F2 x F2 (Самооплодотворение):
Позволим трем генотипическим группам из F2 размножаться отдельно.
F3 (Третье поколение):
- Потомство от DD (из F2) -> 100% правозакрученные.
- Потомство от Dd (из F2) -> 100% правозакрученные.
- Потомство от dd (из F2) -> 100% левозакрученные.
Объяснение: Наконец-то проявляется рецессивный признак. Материнские особи F2 с генотипом dd не имеют доминантного аллеля D, поэтому их яйцеклетки программируются на "левое" развитие.
Скрещивание 2: Левозакрученная мать (Реципрокное)
P (Родители):
- Мать (♀): dd (фенотип: левозакрученная)
- Отец (♂): DD (фенотип: правозакрученный)
F1 (Первое поколение):
- Генотип: Все особи Dd.
- Фенотип: Все 100% особей левозакрученные.
Объяснение: Генотип F1 идентичен Скрещиванию 1 (Dd), но фенотип совершенно другой! Он полностью определяется генотипом матери dd.
F1 x F1 (Самооплодотворение):
Позволяем особям F1 (Dd) размножаться. (Заметьте, эти особи левозакрученные, но несут правозакрученный аллель D).
F2 (Второе поколение):
- Генотипы: 1 DD : 2 Dd : 1 dd.
- Реальный фенотип F2: Все 100% особей — правозакрученные.
Объяснение: Это самый показательный момент. Мать F1 (Dd), хоть и была левозакрученной, имела аллель D. Этот аллель сработал при формировании ее яйцеклеток, запрограммировав ее потомство F2 на правое закручивание. Фенотип потомка определяется генотипом матери, а не фенотипом матери.
F2 x F2 (Самооплодотворение) -> F3 (Третье поколение):
Результат будет абсолютно идентичен Скрещиванию 1:
- Потомство от DD (из F2) -> 100% правозакрученные.
- Потомство от Dd (из F2) -> 100% правозакрученные.
- Потомство от dd (из F2) -> 100% левозакрученные.
"Менделевское" расщепление по фенотипу (3:1) появляется только в поколении F3, с отставанием на одно поколение.
Разгадка: Материнский эффект
Этот феномен "отложенного" наследования объясняется материнским эффектом.
Материнский эффект — это тип наследования, при котором фенотип потомка определяется не собственным генотипом, а генотипом матери. Важно не путать это с цитоплазматическим наследованием (связанным с генами митохондрий или хлоропластов); здесь речь идет о ядерных генах матери.
Механизм:
Направление закрученности раковины определяется на самой ранней стадии развития — во время первых делений дробления зиготы.
- Формирование ооцита (Оогенез): Еще до оплодотворения, в процессе созревания яйцеклетки, материнский организм (в зависимости от своего генотипа) активно синтезирует и "накачивает" будущую яйцеклетку различными веществами: матричной РНК (мРНК), белками и другими регуляторными молекулами. Эти продукты откладываются в цитоплазме ооцита "про запас".
- Активация генома зиготы (ZGA): После оплодотворения собственный геном эмбриона (зиготы) "молчит" в течение первых нескольких делений. Развитие идет исключительно на материнских запасах.
- Ориентация веретена деления: Решающее событие — третье дробление (переход от 4 к 8 клеткам). Материнские продукты (белки) контролируют ориентацию митотических веретен деления.
- Спиральное дробление:
Если материнский белок (продукт аллеля D) присутствует, веретена наклоняются так, что верхние 4 клетки (микромеры) смещаются по часовой стрелке (вправо) относительно нижних. Это декстральное (правое) дробление.
Если этого белка нет (у матери генотип dd), веретена ориентируются иначе, и микромеры смещаются против часовой стрелки (влево). Это синистральное (левое) дробление. - Необратимость: Это решение, принятое на стадии 8 клеток, необратимо. Оно задает асимметрию всего дальнейшего развития, расположение органов и, как следствие, закрученность раковины.
- "Запоздалое" включение: Собственные гены эмбриона (например, аллель d у особи Dd) активируются после того, как это решение уже принято. Они слишком поздно "включаются в работу" и не могут повлиять на уже заданное направление дробления.
Молекулярная основа и современный взгляд
Современная биология развития (Evo-Devo) и генетика подтвердили эту гипотезу. Долгое время ген оставался неизвестным, но в 2019 году ген, ответственный за хиральность у Lymnaea stagnalis, был окончательно идентифицирован — это ген Lsdia1, кодирующий белок формин.
Формины — это белки-организаторы, которые управляют сборкой актинового цитоскелета. Актиновый цитоскелет — это динамичная сеть белковых нитей (микрофиламентов) внутри клетки, ее "силовой каркас" и "рельсы" для внутриклеточного транспорта. Именно он отвечает за физическую ориентацию веретена деления во время митоза.
- Доминантный аллель D (Lsdia1): Кодирует функциональный белок формин. Этот белок (синтезированный матерью и запасенный в яйцеклетке) правильно организует цитоскелет, что приводит к декстральному (правому) дроблению.
- Рецессивный аллель d: Является loss-of-function мутацией (мутацией с потерей функции). Он кодирует нефункциональный белок (или не кодирует его вовсе). В отсутствие функционального формина, организация цитоскелета происходит по-другому, что приводит к синистральному (левому) дроблению.
Доказательство было получено с помощью технологии CRISPR-Cas9. Ученые взяли улиток чистой "правой" линии (DD) и "выключили" (нокаутировали) у них обе копии гена Lsdia1. Как и предсказывала теория материнского эффекта, сами эти улитки остались правозакрученными. Однако все их потомство (поколение F1) оказалось левозакрученным, что и подтвердило роль этого гена как главного "переключателя" хиральности.
Заключение и эволюционное значение
Наследование закрученности раковины у прудовика — это хрестоматийный пример того, как генетика и биология развития (эмбриология) неразрывно связаны. Он показывает, что фенотип не всегда является прямым отражением собственного генотипа, но может определяться "запрограммированным" вкладом материнской клетки.
Эволюционное значение этого механизма огромно. Левозакрученная особь (мутант dd) в правозакрученной популяции (DD) практически не имеет шансов оставить потомство. Из-за "зеркального" строения тела (включая половые органы) спаривание между декстральными и синистральными особями физически невозможно или крайне затруднено.
Таким образом, мутация в одном-единственном гене (D -> d), контролируемом материнским эффектом, выступает как мощнейший механизм мгновенной репродуктивной изоляции. Это один из самых ярких примеров того, как видообразование (в данном случае, симпатрическое, без географической изоляции) может быть запущено изменением в одном гене, отвечающем за раннее развитие.
Список использованной литературы
- Gilbert, S. F. (2010). Developmental Biology. 9th ed. Sunderland, MA: Sinauer Associates.
- Griffiths, A. J. F., Wessler, S. R., Carroll, S. B., & Doebley, J. (2012). Introduction to Genetic Analysis. 10th ed. New York: W. H. Freeman.
- Kuroda, R., Endo, B., Abe, M., & Shimizu, M. (2019). The development of CRISPR for a mollusc establishes the formin Lsdia1 as the long-sought gene for snail dextral/sinistral coiling. Development, 146(9), dev175976.
- Sturtevant, A. H. (1923). Inheritance of direction of coiling in Limnaea. Science, 58(1501), 269-270.
- Инге-Вечтомов С.Г. Генетика с основами селекции. — СПб.: Издательство Н-Л, 2010. — 718 с.