CAM-фотосинтез (от англ. Crassulacean Acid Metabolism — кислотный метаболизм толстянковых) — это высокоспециализированный тип фотосинтеза, представляющий собой сложную биохимическую и физиологическую адаптацию к существованию в условиях жесткого дефицита влаги или нерегулярного водоснабжения. Впервые он был детально изучен у растений семейства Толстянковые (Crassulaceae), откуда и получил свое название. Растения, использующие этот путь, называют CAM-растениями. Этот механизм является ярким примером конвергентной эволюции, поскольку он независимо возникал во многих неродственных группах растений, столкнувшихся с проблемой выживания в засушливых условиях.

Фундаментальная дилемма: Газообмен против испарения
Для любого наземного растения существует фундаментальный физиологический конфликт, который становится особенно острым в аридном климате:
- Потребность в CO₂: Для синтеза органических веществ в процессе фотосинтеза необходим постоянный приток углекислого газа из атмосферы.
- Механизм поглощения: CO₂ поступает в лист через устьица — микроскопические поры, образованные двумя замыкающими клетками. Когда устьица открыты, газообмен возможен.
- Неизбежная плата — транспирация: Движущей силой транспирации (испарения воды) является градиент водного потенциала между насыщенной водяным паром внутренней средой листа и гораздо более сухим окружающим воздухом. Открытие устьиц для поглощения CO₂ неизбежно создает путь для выхода водяного пара.
В жарком и сухом климате (пустыни, полупустыни) этот градиент огромен. Если растение откроет устьица днем, когда фотосинтез наиболее активен, оно столкнется с катастрофической потерей воды, что приведет к обезвоживанию и гибели. Если же оно будет держать устьица постоянно закрытыми для сохранения влаги, то не сможет получить CO₂ и погибнет от углеродного голодания. CAM-фотосинтез — это элегантное эволюционное решение, позволяющее обойти эту дилемму.
Ключевой принцип: Разделение процессов во времени
Если C₄-растения разделили этапы фиксации CO₂ в пространстве (между клетками мезофилла и обкладки проводящего пучка), то CAM-растения пошли по другому пути: они разделили их во времени — между прохладной влажной ночью и жарким сухим днем. Весь процесс происходит в пределах одной и той же клетки мезофилла, но в разные фазы суточного цикла.
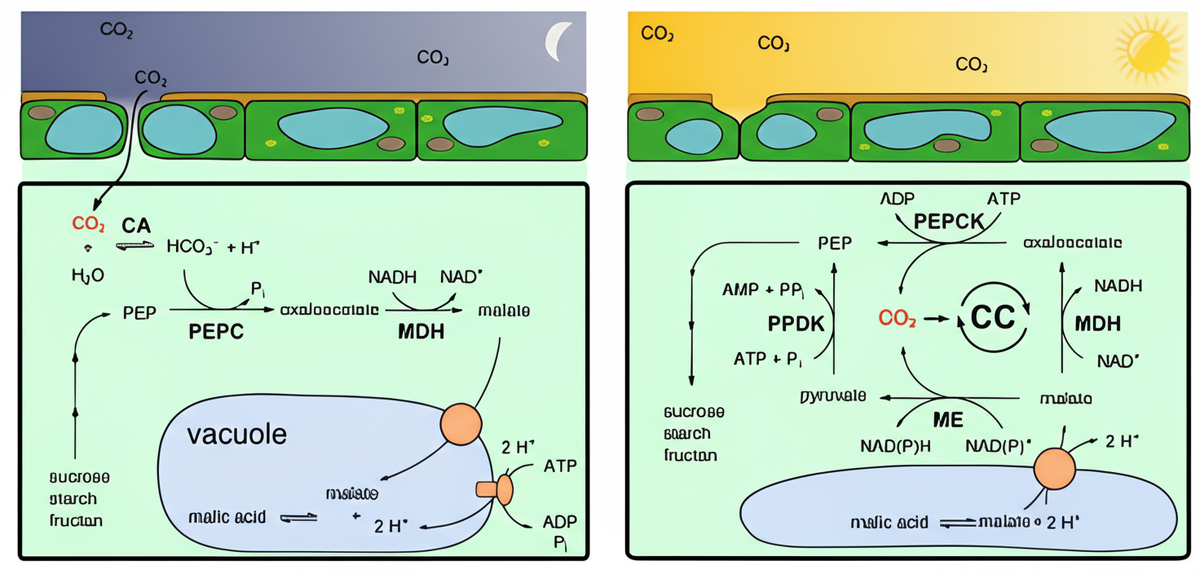
Фаза 1: Ночь (Карбоксилирование — Фиксация CO₂)
Ночью, когда температура воздуха падает, а относительная влажность повышается, градиент водного потенциала между листом и атмосферой минимален. Это "безопасное окно" для газообмена.
- Устьица открываются. Растение начинает активно поглощать CO₂ из атмосферы. Потери воды при этом в десятки раз меньше, чем были бы днем.
- Первичная фиксация CO₂. Попав в цитоплазму, CO₂ с помощью фермента карбоангидразы гидратируется до бикарбонат-иона (HCO₃⁻). Именно эта форма связывается с фосфоенолпируватом (ФЕП) при помощи фермента ФЕП-карбоксилазы. Этот фермент обладает высоким сродством к HCO₃⁻ и, в отличие от Рубиско, не реагирует с кислородом, что делает фиксацию очень эффективной.
- Образование и накопление органических кислот. В результате образуется четырехуглеродная щавелевоуксусная кислота, которая немедленно восстанавливается ферментом малатдегидрогеназой до яблочной кислоты (малата).
- Накопление в вакуоли. Яблочная кислота активно закачивается через мембрану вакуоли (тонопласт) внутрь. Этот процесс (называемый "ловушкой для малата") требует энергии и осуществляется специальными белками-транспортерами. В течение ночи в вакуоли накапливается огромное количество яблочной кислоты, что приводит к двум важным последствиям:
Резкое подкисление клеточного сока (pH может падать с ~5.5 до 4.0 и ниже).
Значительное повышение осмотического давления в клетке, что помогает растению поглощать воду из сухой почвы.
Фаза 2: День (Декарбоксилирование и Цикл Кальвина)
С наступлением дня, когда солнце начинает нагревать воздух и влажность падает, растение переходит в режим тотального водосбережения.
- Устьица плотно закрываются. Это полностью предотвращает транспирацию в самый жаркий период. Газообмен с атмосферой прекращается.
- Высвобождение CO₂ из «хранилища». Яблочная кислота транспортируется из вакуоли обратно в цитоплазму.
- Декарбоксилирование. Специальные ферменты (например, НАДФ-малик-энзим) отщепляют от яблочной кислоты CO₂. Этот процесс создает внутри клетки чрезвычайно высокую, "неестественную" концентрацию углекислого газа, которая может в 10-20 раз превышать его концентрацию в атмосфере.
- Вторичная фиксация CO₂ в цикле Кальвина. Этот концентрированный CO₂ поступает в строму хлоропластов, где его фиксирует фермент Рубиско. Благодаря высокому соотношению CO₂/O₂ в окружении Рубиско, его оксигеназная (нежелательная) активность практически полностью подавляется. CO₂ эффективно включается в цикл Кальвина для синтеза углеводов (глюкозы).
- Регенерация акцептора. Оставшаяся после отщепления CO₂ трехуглеродная молекула (пируват) транспортируется в хлоропласт и используется для регенерации ФЕП (с затратой АТФ), который будет ждать следующей ночи для фиксации новой порции CO₂.
Преимущества CAM-фотосинтеза
- Феноменальная водоэффективность. Ключевое преимущество. Эффективность использования воды (WUE) у CAM-растений поразительна. Если C₃-растения теряют 500-1000 г воды на 1 г фиксированного CO₂, C₄-растения — 250-300 г, то CAM-растения — всего 50-100 г. Это позволяет им не просто выживать, а доминировать в экстремально засушливых местообитаниях.
- Полное подавление фотодыхания. Дневной механизм "суперконцентрации" CO₂ вокруг Рубиско полностью исключает неэффективный и расточительный процесс фотодыхания, повышая чистый выход фотосинтеза в условиях, когда он вообще был бы невозможен.
- Пластичность (Факультативный CAM). Некоторые растения (например, Mesembryanthemum crystallinum) в нормальных условиях фотосинтезируют по C₃-пути, но при наступлении засухи или засоления способны "включать" CAM-механизм. Это демонстрирует невероятную адаптивную гибкость данной метаболической стратегии.
Недостатком является более медленный общий темп роста по сравнению с C₃- и C₄-растениями в благоприятных условиях, так как емкость вакуоли для накопления кислоты и количество энергии, затрачиваемой на транспорт и регенерацию, ограничены.
Сравнительная таблица путей фотосинтеза
Примеры CAM-растений
- Суккуленты: Подавляющее большинство пустынных растений. Их морфологическая особенность — наличие мясистых, водозапасающих тканей (суккулентность) — идеально сочетается с биохимией CAM-фотосинтеза.
Семейство Кактусовые (Cactaceae)
Семейство Толстянковые (Crassulaceae) — очитки, каланхоэ.
Род Алоэ (Aloe)
Род Агава (Agave) - Эпифиты: Растения, живущие на стволах и ветвях других растений. Несмотря на высокую влажность воздуха в тропических лесах, они испытывают «физиологическую сухость», так как их корни не контактируют с почвой, и они могут использовать только ту воду, что выпадает с дождем.
Многие Орхидеи (Orchidaceae)
Бромелиевые (Bromeliaceae), включая ананас (Ananas comosus). - Некоторые водные растения: Например, полушник (Isoetes). Для них CAM — это способ запасать CO₂ ночью, когда его концентрация в воде выше из-за дыхания других организмов, и использовать днем для фотосинтеза.
Список рекомендуемой литературы
- Sage, R. F. (2016). A portrait of the C₄ photosynthetic family on the 50th anniversary of its discovery: species number, evolutionary lineages, andHall of Fame. Journal of Experimental Botany, 67(14), 4039-4056. (Хотя статья посвящена C₄-пути, в ней проводятся важные сравнения с CAM-фотосинтезом).
- Winter, K., & Smith, J. A. C. (Eds.). (1996). Crassulacean Acid Metabolism: Biochemistry, Ecophysiology and Evolution. Springer-Verlag. (Фундаментальная монография, полностью посвященная CAM-фотосинтезу).
- Taiz, L., Zeiger, E., Møller, I. M., & Murphy, A. (2015). Plant Physiology and Development (6th ed.). Sinauer Associates. (Глава «Photosynthesis: Carbon Reactions» содержит детальное и современное описание всех трех путей фотосинтеза).
- Ермаков И. П. (ред.) (2005). Физиология растений. М.: Академия. (Качественный отечественный учебник для вузов).