Данное пособие предлагает углубленный разбор кровеносной системы костистых рыб (Teleostei). Мы отходим от базового ЕГЭ-шного описания "одного круга и двухкамерного сердца" и фокусируемся на специфических анатомических и физиологических адаптациях, которые критически важны для понимания этой группы на олимпиадном уровне.
Физические и Физиологические Ограничения
Кровеносная система рыб — это прямое следствие физики водной среды.
- Низкая растворимость O₂: Вода (полярный растворитель) плохо растворяет неполярный O₂. Концентрация кислорода в воде в 20-30 раз ниже, чем в воздухе. Это требует от органа дыхания (жабр) максимально возможной эффективности извлечения, что достигается огромной площадью поверхности и противоточным механизмом газообмена. Как мы увидим, эта эффективность имеет высокую гемодинамическую "цену".
- Высокая плотность воды: Вода в ~800 раз плотнее воздуха. Это обеспечивает нейтральную плавучесть (поддержку тела). В результате сердце рыбы освобождается от задачи преодоления силы тяжести. Ему не нужно создавать высокое давление, чтобы поднять кровь к мозгу, как у жирафа. Это ключевой фактор, позволивший системе работать при низком давлении.
- Пойкилотермия: Метаболизм рыб напрямую зависит от температуры среды (описывается коэффициентом Q10, часто ~2-3, означая удвоение-утроение скорости метаболизма при повышении t° на 10°C). Это означает, что потребность в O₂ непостоянна. Система должна справляться с пиковыми нагрузками (охота), но большую часть времени работает в экономичном режиме.
Главная задача: Пропустить 100% сердечного выброса через высокоэффективную (и создающую колоссальное сопротивление) капиллярную сеть жабр, а затем, без дополнительного насоса, доставить эту кровь к тканям.
Анатомия Сердца
Сердце костистых рыб представляет собой насос, состоящий из четырех последовательных отделов. Его фундаментальная особенность заключается в том, что оно пропускает через себя исключительно деоксигенированную кровь, которую получает от тканей тела и направляет к жабрам.
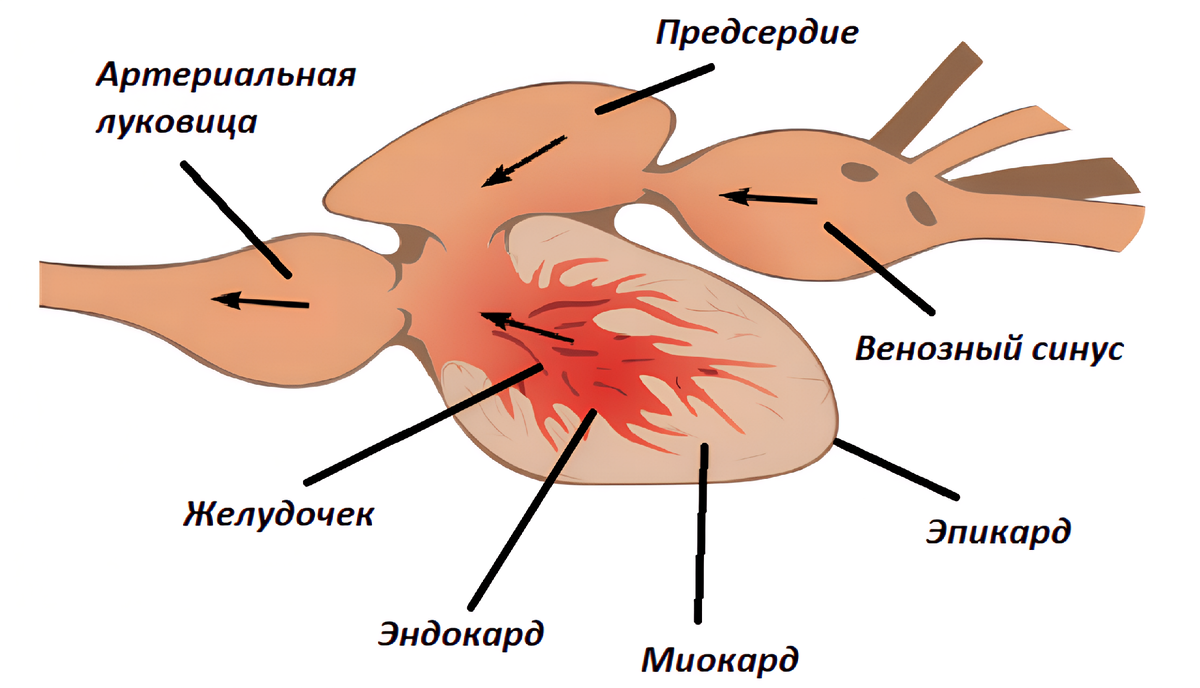
- Венозный синус (Sinus Venosus): Тонкостенный коллектор, являющийся истинной первой камерой. Он принимает кровь из Кювьеровых протоков (объединенные передние и задние кардинальные вены) и печеночной вены. Его стенки содержат кардиомиоциты и являются главным водителем ритма (гомолог синоатриального узла млекопитающих).
- Предсердие (Atrium): Более мускулистый отдел, работающий как "насос предварительной подкачки" (booster pump). Его сокращение ("предсердный толчок") активно наполняет желудочек, растягивая его миокард и обеспечивая более сильное последующее сокращение по механизму Франка-Старлинга.
- Желудочек (Ventriculus): Основной силовой насос. Толстостенный, мышечный, часто пирамидальной или мешковидной формы. Его мощная систола создает пиковое давление (50-70 мм рт. ст.) для всего круга. Его относительная масса сильно варьирует: от <0.1% массы тела у малоподвижных рыб до >0.5% у активных тунцов.
- Сравнительный аспект: Луковица vs. Конус: Это фундаментальный таксономический и функциональный признак.
- У Костистых рыб (Teleostei): Луковица Аорты (Bulbus Arteriosus). Это толстостенное, эластичное образование, производное брюшной аорты. Состоит из эластина и гладких мышц, в нем нет кардиомиоцитов и нет клапанов. Его функция — пассивно-эластическая.
- У Хрящевых рыб (Chondrichthyes) и Двоякодышащих: Артериальный Конус (Conus Arteriosus). Это мышечная, сократимая камера сердца (производное сердечной трубки). Содержит кардиомиоциты и несколько рядов клапанов. Он активно сокращается, помогая желудочку и дополнительно повышая давление.
Клапанный Аппарат
Для предотвращения ретроградного (обратного) тока крови в сердце имеется строгая система пассивных клапанов, работающих по градиенту давления.
- Синоатриальный клапан (S-A Valve): На границе венозного синуса и предсердия. Обычно двустворчатый, мембранный. Открывается при расслаблении предсердия, закрывается при его систоле.
- Атриовентрикулярный клапан (A-V Valve): Между предсердием и желудочком. Крупный, как правило, двустворчатый. Пассивно захлопывается под давлением во время систолы желудочка.
- Клапаны на выходе из желудочка: У костистых рыб на границе желудочка и луковицы аорты есть пара полулунных клапанов. Они предотвращают обратный ток из эластичной луковицы (которая находится под давлением) в расслабляющийся желудочек (во время диастолы).
Механика Насоса и "Аспирационное Наполнение" (Vis-a-fronte):
Сердце рыб находится в ригидном (жестком) перикарде.
- Систола желудочка: Желудочек выбрасывает кровь. Из-за своей пирамидальной формы его объем внутри перикарда резко уменьшается.
- Эффект: Это создает отрицательное давление (вакуум, ~ -1 до -3 мм рт. ст.) внутри жесткого перикарда.
- Результат: Это отрицательное давление "растягивает" тонкостенные венозный синус и предсердие, буквально "засасывая" (аспирируя) в них венозную кровь из вен, где давление почти нулевое.
- Значение: Это активный механизм "vis-a-fronte" (тяга спереди), который обеспечивает венозный возврат к сердцу, в отличие от наземных позвоночных, которые полагаются на "vis-a-tergo" (давление сзади от венозного русла). Этот механизм возможен только в условиях нейтральной плавучести.
Физиология Миокарда
Ключевой олимпиадный вопрос: как сердце, качающее 100% венозной крови, само получает кислород?
- Тип I: Губчатый (Трабекулярный) Миокард: У большинства костистых рыб (особенно у малоподвижных, донных) желудочек имеет губчатую, трабекулярную структуру. Он не имеет коронарной системы. Кардиомиоциты получают O₂ и питательные вещества путем диффузии прямо из венозной крови, протекающей через камеру.
- Ограничение: Это серьезный лимит. Кровь в желудочке — самая деоксигенированная во всем теле. При физической нагрузке (когда ткани забирают больше O₂) сердце оказывается на грани гипоксии. Это главный фактор, ограничивающий пиковую и длительную активность.
- Тип II: Компактный + Губчатый Миокард: У активных, быстрых пелагических рыб (например, тунец, лосось, макрель) желудочек имеет два слоя: внутренний губчатый и мощный наружный компактный слой. Этот компактный слой имеет собственную коронарную систему.
- Нюанс коронарной системы: Коронарные артерии отходят ПОСЛЕ жабр (например, от выносящих жаберных артерий или от спинной аорты). Они приносят к миокарду артериальную кровь, но под низким давлением. Это пример конвергентной эволюции с амниотами и ключевая адаптация, позволившая тунцам развить элементы эндотермии ("теплокровности").
Гемодинамика
Гемодинамика — это история о давлении.
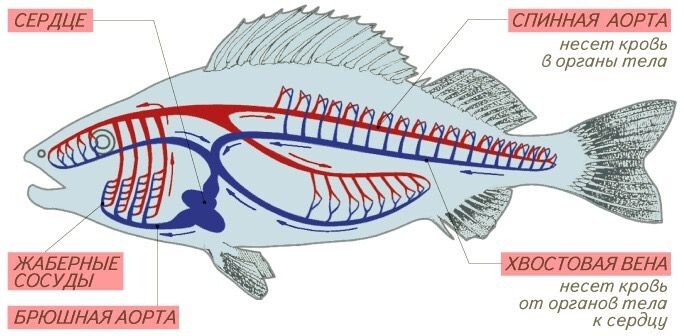
- Желудочек -> Брюшная Аорта: Давление высокое, пульсирующее (пик ~50-70 мм рт. ст.).
- Луковица Аорты: Эластично растягивается, демпфирует (сглаживает) пиковое давление и превращает пульсирующий поток в ламинарный (постоянный). Это защищает нежные жабры и повышает эффективность газообмена. Давление на входе в жабры ~40-60 мм рт. ст.
- Капилляры Жабр (1-я сеть): Это зона наибольшего сопротивления. Согласно закону Пуазейля, сопротивление обратно пропорционально радиусу в 4-й степени (R ∝ 1/r⁴). Миллиарды тончайших капилляров, необходимых для диффузии, создают огромное суммарное сопротивление.
- Результат: При прохождении этой сети теряется ~80-90% давления.
- Выносящие жаберные артерии -> Спинная Аорта: Кровь артериальная, но давление в спинной аорте критически низкое (~10-15 мм рт. ст.).
- Капилляры Тела (2-я сеть): Давление падает еще ниже (~5-10 мм рт. ст.), едва обеспечивая перфузию (орошение) тканей. Кровоток в теле медленный.
- Венозная система: Давление близко к нулю. Возврат крови к сердцу обеспечивается мышечными сокращениями ("мышечная помпа") и, главное, механизмом аспирационного наполнения.
Эволюционное Значение
Система костистых рыб — это высокоспециализированный, но компромиссный механизм.
- Компромисс: Максимальная эффективность газообмена в жабрах была "куплена" ценой хронически низкого давления (гипотензии) в большом круге кровообращения.
- Лимит: Эта система не может обеспечить высокий метаболизм (теплокровность, за исключением адаптаций тунцов) или эффективную борьбу с гравитацией (жизнь на суше).
- Эволюционный Вектор: Единственным решением этой проблемы стало разделение кругов кровообращения. Это позволило после органа дыхания (легких) повторно направить кровь в сердце (в левое предсердие) и создать высокое давление в системном круге, не повреждая при этом нежные легочные капилляры.
- Переход: Этот переход начинается у Двоякодышащих рыб (Dipnoi), у которых в ответ на появление легочного дыхания развивается неполная межпредсердная перегородка и способность частично разделять потоки, направляя артериальную кровь от легких обратно в сердце. Это прямой эволюционный "эскиз" системы всех четвероногих (Tetrapoda).
Список использованной литературы
- Kardong, K. V. (2018). Vertebrates: Comparative Anatomy, Function, Evolution. 8th ed. McGraw-Hill. (Chapter 12: Circulatory System).
- Randall, D., Burggren, W., French, K. (2002). Eckert Animal Physiology: Mechanisms and Adaptations. 5th ed. W.H. Freeman. (Chapter 12: Circulation).
- Farrell, A. P. (2011). Encyclopedia of Fish Physiology: From Genome to Environment. Academic Press. (Section IV: Cardiovascular System).
- Дзержинский Ф. Я., Васильев Б. Д., Малахов В. В. (2013). Зоология позвоночных. 2-е изд. "Академия".
- Liem, K. F., Bemis, W. E., Walker, W. F., & Grande, L. (2001). Functional Anatomy of the Vertebrates: An Evolutionary Perspective. 3rd ed. Harcourt College Publishers.
- Burggren, W. (2013). Comparative Animal Physiology. Cambridge University Press.