Данное пособие посвящено плавательному пузырю — органу, который часто ошибочно считают уникальной "рыбьей" структурой. На самом деле, это одна из двух версий органа, изначально возникшего для дыхания воздухом. Понимание плавательного пузыря — это ключ к пониманию того, как возникли наши собственные легкие.
Эволюционная Дилемма: Дыхание или Плавучесть?
И плавательный пузырь (у Лучеперых рыб, Actinopterygii), и легкие (у Лопастеперых, Sarcopterygii, и у нас) являются гомологичными структурами. Они оба развились как дивертикулы (выросты) эмбриональной пищеварительной трубки в девонском периоде ("Веке Рыб").
Первичные костные рыбы (Osteichthyes) столкнулись с двумя фундаментальными проблемами, требовавшими решения:
- Проблема 1 (Дыхание): Девонские пресные водоемы были теплыми, часто застойными и, как следствие, бедными кислородом (гипоксия). Одних только жабр для выживания в таких условиях было недостаточно. Требовался вспомогательный орган для дыхания атмосферным воздухом.
- Проблема 2 (Физика Плавучести): Плотность костей (~2.0 г/см³) и мышечной ткани (~1.06 г/см³) выше плотности воды (~1.0 г/см³). Без вспомогательных механизмов костная рыба тонет.
Решение Хрящевых рыб (Акул): Акулы решили эту проблему "метаболически дорого": они накапливают в печени легкий жир (сквален, ~0.86 г/см³) и постоянно поддерживают гидродинамическую подъемную силу за счет движения.
Решение Костистых рыб: Нужен был более эффективный механизм — внутренний "поплавок", позволяющий достичь нейтральной плавучести и экономить энергию.
Эволюция разделила костных рыб на два пути, основанных на том, как расположился этот вырост:
- Путь 1 (Легкие): У наших предков (Лопастеперых) вырост пошел вниз (вентрально). Это анатомически выгодно для дыхания (легко дренировать слизь), но создает гидростатическую неустойчивость (центр плавучести ниже центра тяжести, что переворачивает рыбу). Орган специализировался на дыхании.
- Путь 2 (Пузырь): У предков Лучеперых рыб вырост пошел вверх (дорсально). Это выгодно для стабильности (центр плавучести над центром тяжести), но невыгодно для дыхания (проблемы с дренажем). Орган специализировался на плавучести.
Модель 1: Физиостомы (Physostomi) — "Открытопузырные"
Это эволюционно более древняя модель, сохранившаяся у таких групп, как лососевые, карповые, сельдевые, угревые и тарпоны.
- Анатомия: Плавательный пузырь сохраняет пожизненную связь с пищеводом через специальный проток — ductus pneumaticus (пневматический проток). Состав газа в пузыре близок к составу воздуха (высокое содержание N₂).
- Механизм Регуляции: Прямой, "грубый", но быстрый.
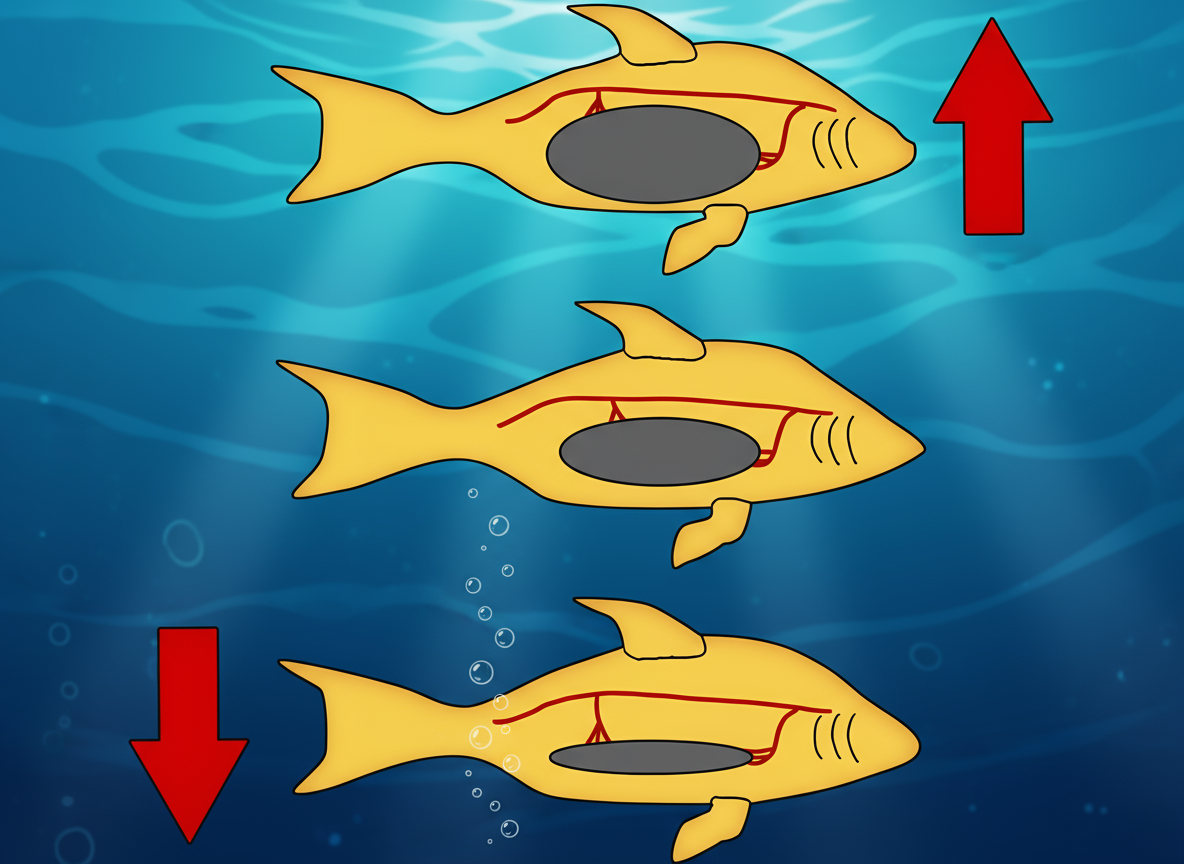
Чтобы уменьшить плавучесть (погрузиться): Рыба сжимает тело и "выпускает" газ через проток, рот и жаберную крышку.
Чтобы увеличить плавучесть (всплыть): Рыба должна физически подняться к поверхности и заглотнуть воздух. - Адаптивное Значение:
Плюс: Быстрая регуляция объема. Позволяет совершать быстрые вертикальные миграции без риска баротравмы.
Минус (Критический): Рыба экологически привязана к поверхности. Она не может долго находиться на большой глубине, не всплывая для "подкачки". Всплытие для глотка воздуха также повышает риск быть съеденным воздушными хищниками.
Эволюционный Компромисс: У многих "продвинутых" физиостом (например, у карпа или угря) уже есть примитивные газовые железы и Rete Mirabile. Они могут секретировать газ, но основной и самый быстрый механизм — все еще заглатывание.
Модель 2: Физиоклисты (Physoclisti) — "Закрытопузырные"
Это эволюционно "продвинутая" модель, характерная для большинства современных костистых рыб (окунеобразные, трескообразные и т.д.). Она позволила рыбам полностью "отвязаться" от поверхности и освоить любые глубины.
- Анатомия: Пневматический проток (ductus pneumaticus) полностью редуцируется (зарарастает) в ходе развития. Пузырь становится полностью изолированным мешком, наполненным газом (преимущественно O₂).
- Механизм Регуляции: Вместо "заглатывания" у них развились две сложные биохимические "машины" для накачки и сброса газа.
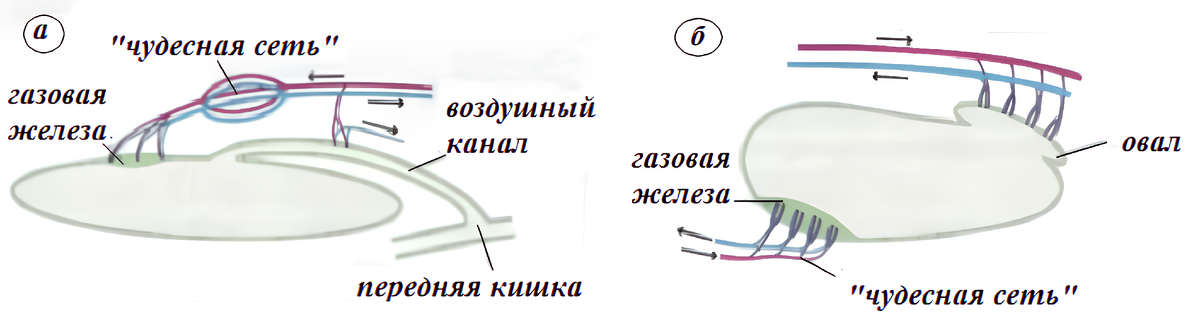
А. Накачка Газа: Газовая Железа и "Чудесная Сеть" (Rete Mirabile)
Это один из самых поразительных примеров противоточного множителя в биологии, способный накачивать газ против колоссального градиента давления (на глубине 1000 м давление в 100 раз выше, чем на поверхности).
- Газовая Железа (Gas Gland): Участок стенки пузыря. Его клетки при стимуляции (сигнал от нервной системы) переключаются на интенсивный анаэробный гликолиз (даже в присутствии кислорода!). Они производят и выделяют в кровь молочную кислоту и CO₂.
- Эффекты в Крови (Закисление): Молочная кислота (H⁺) вызывает два ключевых эффекта в гемоглобине рыб:
Эффект Бора: При ↓pH (закислении) сродство гемоглобина к O₂ снижается (кривая диссоциации сдвигается вправо). Гемоглобин легче отдает O₂.
Эффект Рута: (Ключевой!) При ↓pH гемоглобин рыб снижает свою максимальную кислородную емкость. Он не просто легче отдает O₂, он физически не может удержать то же количество O₂, даже при высоком парциальном давлении. - Результат: Кровь, проходящая через газовую железу, насильно "выталкивает" из гемоглобина растворенный O₂. Парциальное давление p(O₂) в плазме этой крови резко возрастает.
- Rete Mirabile ("Чудесная Сеть"): Это плотный пучок из тысяч артериальных и венозных капилляров, идущих параллельно друг другу (противотоком).
Кровь, выходящая от железы (венозный капилляр), имеет аномально высокое p(O₂).
Растворенный O₂ начинает диффундировать по градиенту... прямо в соседний артериальный капилляр, идущий к железе.
Механизм "Ловушки" (Противоточный Обменник): Кислород "запирается" в этой петле. Он циркулирует и накапливается в сети, не уходя в общий кровоток.
Механизм "Множителя": С каждым циклом концентрация O₂ в сети растет (умножается). p(O₂) в Rete Mirabile становится в десятки и сотни раз выше, чем в артериальной крови.
Когда p(O₂) в "чудесной сети" становится выше, чем p(O₂) внутри пузыря, O₂ начинает диффундировать внутрь пузыря, накачивая его.
Б. Сброс Газа: "Овал" (Oval)
Для сброса газа (при всплытии) используется другой механизм.
- Анатомия: В другом конце пузыря есть участок ("Овал"), богатый "обычными" капиллярами, который изолирован от пузыря мышцей-сфинктером.
- Механизм: Когда рыбе нужно уменьшить плавучесть, сфинктер расслабляется. Газ из пузыря (где p(O₂) сверхвысокое) начинает диффундировать в кровь "Овала" (где p(O₂) низкое, как в обычной венозной крови) и уносится в общий кровоток, выводясь затем через жабры.
Эволюционное и Адаптивное Значение
- Нейтральная Плавучесть: Главная функция. Освободила рыб от необходимости тратить энергию на постоянное плавание, чтобы не утонуть (как акулы) или не всплыть. Экономия энергии колоссальна.
- Освоение Глубин: Модель Физиоклист позволила рыбам освоить все океанические глубины, полностью разорвав связь с поверхностью.
- Ограничение (Баротравма): Система физиоклист медленная (и накачка, и сброс). При быстром подъеме (например, когда рыбу ловит рыбак) газ не успевает сброситься через Овал. По Закону Бойля (P₁V₁=P₂V₂), давление падает, а объем газа в пузыре катастрофически растет, раздувая и убивая рыбу.
- Вторичные Функции (Ко-опция):
Дыхание: У некоторых "примитивных" физиостом (амия, панцирник) пузырь сохранил густую сеть капилляров и используется как вспомогательное легкое в бедной O₂ воде.
Слух (Веберов аппарат): У карпообразных, сомов и харациновых (Otocephala) развился Веберов аппарат — система из 4-х парных косточек (tripus, intercalarium, scaphium, claustrum), которая механически соединяет стенку пузыря с внутренним ухом. Пузырь работает как резонатор (усилитель звука), а косточки — как передатчик, резко обостряя слух в сотни раз.
Звукоизвлечение: Некоторые рыбы (например, горбылевые, рыбы-жабы) имеют звуковые мышцы (sonic muscles), прикрепленные к пузырю. Эти мышцы способны к сверхбыстрым сокращениям (самым быстрым среди позвоночных), заставляя пузырь вибрировать и издавать громкие звуки для коммуникации.
Список использованной литературы
- Kardong, K. V. (2018). Vertebrates: Comparative Anatomy, Function, Evolution. 8th ed. (Chapter 11: Digestive System, Chapter 13: Respiratory System).
- Hickman, C. P., Roberts, L. S., et al. (2017). Integrated Principles of Zoology. 17th ed. (Chapter 24: Fishes).
- Liem, K. F., Bemis, W. E., et al. (2001). Functional Anatomy of the Vertebrates: An Evolutionary Perspective.
- Schmidt-Nielsen, K. (1997). Animal Physiology: Adaptation and Environment. 5th ed. (Chapter 5: Buoyancy & Gas Glands).
- Pelster, B. (2015). The generation of high gas partial pressures in fish swim bladders. Comparative Biochemistry and Physiology Part A, 185, 78-87.