Дендритные шипики – малые актин-богатые выросты на дендритах пирамидных нейронов, обеспечивающие локализацию большинства возбуждающих синапсов и их пластичность. Динамическое полимеризование F-актина в шипиках определяет их морфологические изменения и тесно связано с обучением и памятью. Центральный регулятор этой системы – LIM-киназа 1 (LIMK1), фосфорилирующая и инактивирующая актин-связывающий белок кофилин. Фосфорилированный кофилин не расщепляет нити F-актина, что стабилизирует цитоскелет шипика и способствует его расширению. Таким образом, баланс между фосфорилированным (инактивным) и нефосфорилированным (активным) кофилином является ключевым фактором регуляции динамики актина и синаптической пластичности.
Основные механизмы формирования и стабилизации шипиков
Полимеризация актиновых филаментов в шипиках осуществляется преимущественно за счёт ветвящихся нитей, инициированных комплексом Arp2/3. Активатор Arp2/3 – WAVE-регуляторный комплекс (WRC) – содержит субъединицы (Cyfip1, Nckap1, Abi1/2 и др.), критичные для морфогенеза шипиков. Мутации или потеря компонентов WRC приводят к дефициту шипиков и нарушению синаптической пластичности, что в моделях мышей проявляется в когнитивных и поведенческих дефицитах. Например, Sempert et al. (2023) показали, что рецептор Neogenin и его лиганд RGMa активируют WRC‑Arp2/3, стимулируя ветвящую полимеризацию актина в шипиках и их расширение; при дефиците Neogenin/RGMa наблюдается избыток «филоподий» (шупальцевых выростов) и уменьшение числа зрелых «грибовидных» шипиков. Таким образом, модуляторы WRC (Neogenin/RGMa) являются критическими регуляторами структурной пластичности шипиков.
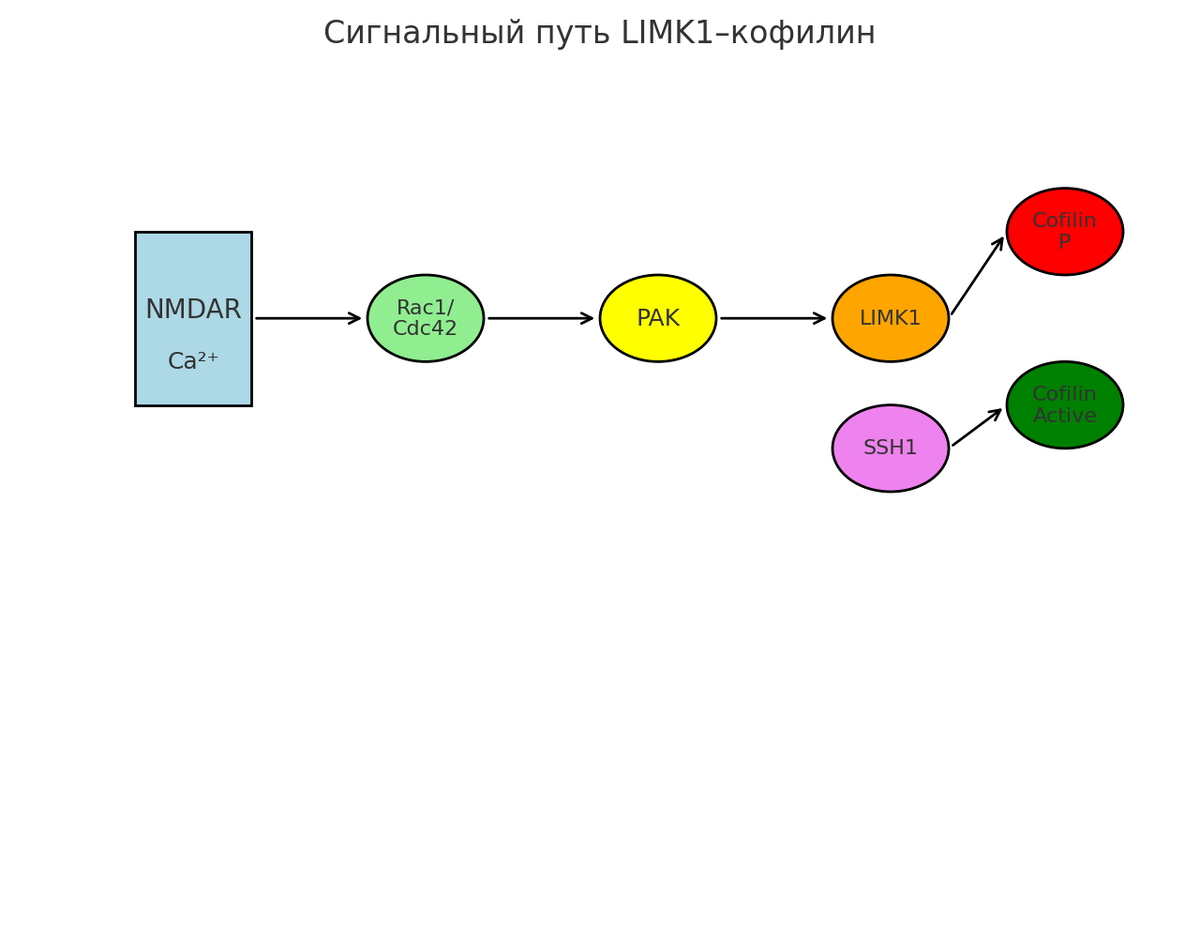
Верхние уровни сигнализации включают Rho-GTPазы (Rac1, Cdc42, RhoA), которые по событиям синаптической активности (например, при Ca^2+‑входе через NMDAR) активируют эффекторные киназы (PAK, ROCK) и, в итоге, LIMK1. Активация LIMK1 ведёт к фосфорилированию кофилина (Ser3) и его инактивированию. Обратно, фосфатаза Slingshot-1 (SSH1) дефосфорилирует кофилин, восстанавливая его активность. Этот цикл фосфорилирования-дефосфорилирования кофилина обеспечивает динамическое чередование стабилизации и ремоделирования F-актина в шипиках.
Кроме того, ряд специальных актин-связывающих белков обеспечивает организацию цитоскелета шипиков. Актин-каппер (CP), например, присоединяется к «барбитовым» (+)-концам нитей актина, препятствуя их удлинению, и способствует образованию плотных ветвящихся сетей. Удаление CP в нейронах снижает число зрелых шипиков и увеличивает долю филаментообразных выростов, указывая на роль CP в стабилизации зрелой формы шипиков. Распределение CP в шипиках зависит от взаимодействия с CPI-мотивными адапторами (например, Twinfilin-1), и нарушает сборку постсинаптического белкового комплекса при дефиците этих белков. Другой пример – профилин2а (PFN2a), связывающий мономеры актина. Фосфорилирование PFN2a в положении Ser137 сильно влияет на базовую динамику F-актина и форму шипиков: мутации, имитирующие де/фосфорилирование PFN2a, снижают диаметр головки шипика и меняют кинетику обмена актина. Известно также, что PFN2a необходим для роста головы шипика при индуцированном LTP, что подчёркивает её роль в активности-зависимой пластичности.
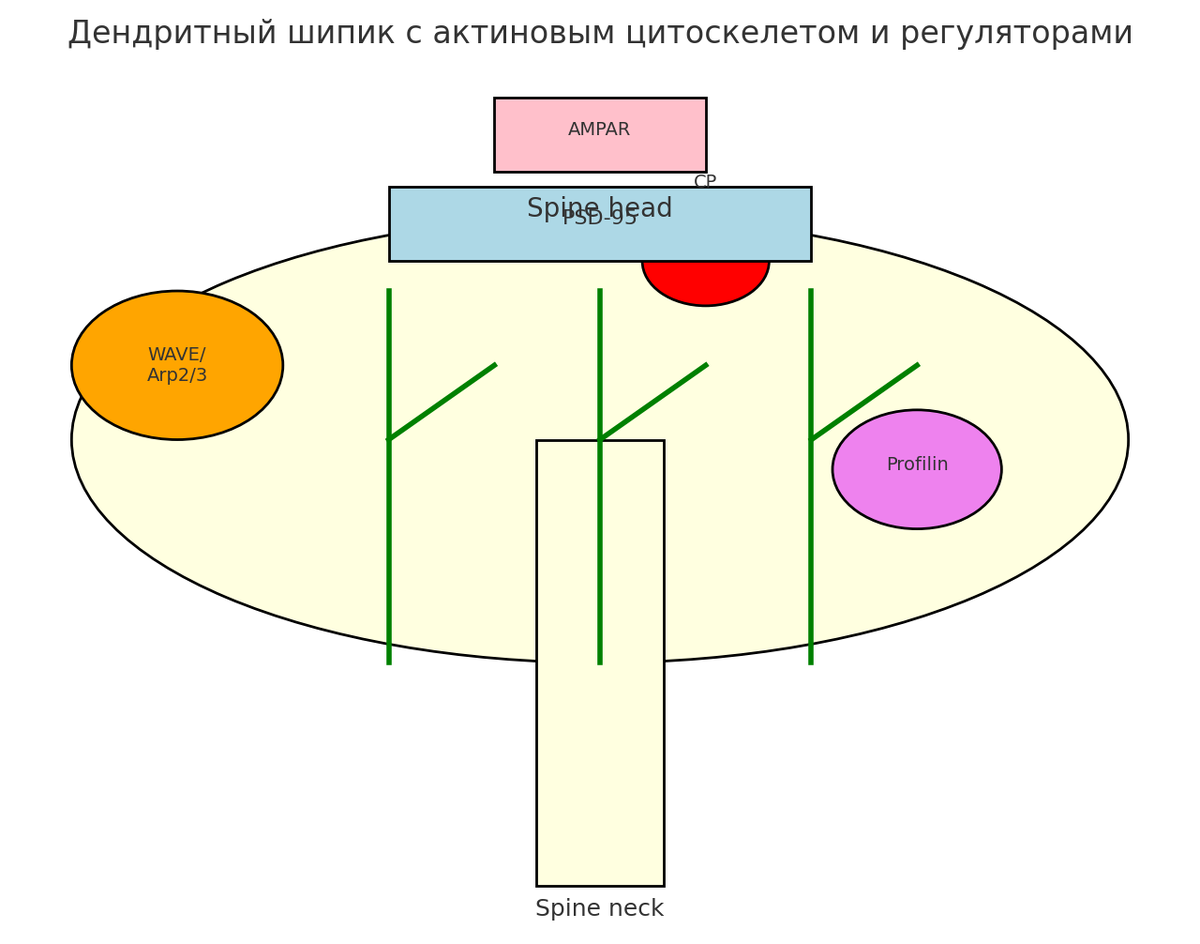
И, наконец, F-актин в шипиках перекрёстно-связан с постсинаптическими белками (например, с дребрином и PSD-95), укрепляя структуру шипиков. В совокупности, эти механизмы (ветвление актина через Arp2/3-WAVE, модификация кофилина через LIMK1/SSH1, регуляция за счёт CP, профилина и др.) обеспечивают тонкий контроль морфологии и стабильности дендритных шипиков.
Связь с когнитивными функциями и памятью
Актин-опосредованная структурная пластичность шипиков является молекулярным субстратом обучения и запоминания. Индукция LTP, коррелят обучения, сопровождается быстрым появлением ветвящихся актиновых нитей в головке шипиков и их увеличением в объёме. Данные Sempert et al. подтверждают, что при LTP происходит привлечение Arp2/3 к постсинаптической плотности, что стимулирует рост шипиков и консолидацию усиленной синаптической связи. Нарушения этих процессов отражаются на поведении. Так, при условной делеции Neogenin у мышей обнаружено отсутствие индуцируемого LTP и выраженный дефицит в восстановлении консолидированного страха (fear memory). Аналогично, Tolias и соавторы (2025) показали, что удаление Rac1-геф Tiam1 из взрослых нейронов гиппокампа усиливает обучение и память: Tiam1^fKO-мыши демонстрируют усиленную условно-страховую и пространственную память при повышенной NMDAR-зависимой пластичности. Молекулярно это связывалось с повышенной передачей NMDAR в синапсах и снижением уровня F-актина в шипиках (что отражает повышенную их «турбулентность»), а также с нарушением эндоцитоза NMDAR. Авторы интерпретируют эти результаты так, что Tiam1 обычно ограничивает пластичность гиппокампа, стабилизируя актиновую сеть шипиков и уменьшая доступность NMDAR; при высокой синаптической активности Tiam1 деградирует, устраняя этот «тормоз».
Дисфункция актинового цитоскелета шипиков также отражается на патологических нарушениях памяти. Например, при болезни Альцгеймера отмечается значительное снижение плотности шипиков в гиппокампе и коре при нарастании амилоид-β и тау-патологии. В таких условиях формируются патологические структуры «кофилин-актиновых стержней» (cofilin-actin rods) и «тел Хирано», свидетельствующие об аномальной стабилизации актина. По данным Paciello et al. (2025), нарушение оси LIMK1–кофилин–актин тесно связано с синаптической дегенерацией в моделях болезни Альцгеймера. В частности, гиперактивация LIMK1 (что усиливает фосфорилирование и инактивацию кофилина) и, напротив, повышенная дефосфорилизация кофилина разными авторами связывались с нейродегенеративными изменениями и дефицитом LTP. Эти данные указывают, что коррекция баланса актина в шипиках может быть перспективным подходом для восстановления функций памяти: так, предлагается рассматривать ингибиторы LIMK1 или активаторы Slingshot как потенциальную терапию для улучшения пластичности и снижения когнитивного дефицита.
Кофилин и его фотомодуляция
Кофилин остаётся центральным узловым белком в регуляции динамики F-актина, поскольку именно он контролирует деполимеризацию и фрагментацию филаментов. В последние годы появились подходы, позволяющие не только фармакологически или генетически, но и оптогенетически управлять активностью кофилина.
Недавние работы (2022–2024 гг.) применили фоточувствительные домены (например, LOV2 или CRY2-CIB1) для создания «оптокофилина» — конструкта, в котором локальная активация или инактивация кофилина индуцируется лазерным облучением в диапазоне синего света. Такой метод позволил с субсекундным и субмикронным разрешением запускать деполимеризацию актиновых нитей в конкретных дендритных шипиках и отслеживать их морфологические изменения в реальном времени.
Результаты показали, что кратковременное локальное включение кофилина приводит к обратимому уменьшению объёма шипика и снижению постсинаптического кальциевого ответа, тогда как его дезактивация способствует восстановлению морфологии и функциональной активности синапса. Важно, что такая точечная модуляция позволила подтвердить причинно-следственную связь между актиновым ремоделированием, индуцированным кофилином, и процессами долговременного потенцирования (LTP).
Фотомодуляция кофилина рассматривается как новый инструмент для прямого тестирования гипотезы о том, что динамика актина в шипиках является не просто коррелятом, а необходимым условием для консолидации памяти. Более того, эти методы позволяют разрабатывать протоколы «лазерной синаптической инженерии», где возможно выборочно ослаблять или усиливать отдельные синапсы. Это открывает путь как к фундаментальному изучению когнитивных процессов, так и к потенциальным терапевтическим стратегиям, например при моделировании патологий, сопровождающихся нарушением актинового цитоскелета (болезнь Альцгеймера, когнитивное старение).
Заключение
Недавние исследования подчёркивают, что динамика актинового цитоскелета в дендритных шипиках – это результат тонкой кооперации множества сигналов и белков. Интеграция Rho-GTPазных путей (Rac1/Cdc42/RhoA – PAK/ROCK – LIMK1/кофилин) с актин-иницирующими комплексами (WAVE/Arp2/3) и регуляторами длины нитей (CP, профилин и др.) определяет формирование, стабилизацию и пластичность шипиков. Эти молекулярные механизмы лежат в основе долговременного потенцирования и когнитивных функций: нарушения их координации приводят к дефицитам памяти (что показано на примере патологий или генетических моделей). Важно отметить, что баланс активности актина (фосфорилированного кофилина vs. активного) является ключевым узлом регуляции синаптической пластичности.
Подробное понимание этих процессов создаёт основу для разработки новых терапевтических стратегий – например, модуляция LIMK1/кофилина или WAVE-активности – направленных на восстановление структурной пластичности шипиков и улучшение когнитивных функций при неврологических заболеваниях. Перспективным направлением также выступает использование методов оптогенетики и лазерной модуляции кофилина, позволяющих с высокой пространственно-временной точностью управлять динамикой F-актина в отдельных шипиках. Такие подходы не только дают возможность прямой проверки гипотезы о связи актинового ремоделирования с памятью, но и открывают путь к разработке новых методов коррекции когнитивных нарушений.
Список ключевых источников:
Paciello F. et al., Cell Death & Disease, 2025
Sempert K. et al., Front. Mol. Neurosci., 2023
Heinze A. & Rust M.B., Eur. J. Cell Biol., 2023
Tolias K.F. et al., J. Neurosci., 2025
Michaelsen-Preusse et al. (PFN2a), Front. Cell Dev. Biol., 2023
Cyclin Y regulates spatial learning and memory flexibility (Seo et al., 2023)
Engineering memory with an extrinsically disordered kinase (обозначается Science Advances, Ripoli et al., 2023)
Z-lock, a Broadly Applicable Optogenetic Method, Controls Cofilin in Living Cells (Stone et al., 2025)