
Фертильность млекопитающих зависит от образования яйцеклетки, способной к оплодотворению и поддерживающей раннее развитие эмбриона. Для этого необходимы как цитоплазматические, так и ядерные, то есть хромосомные, процессы, которые были инициированы за десятилетия до овуляции. Текущие демографические изменения, связанные с задержкой материнства, все чаще вступают в противоречие с этими биологическими процессами.
Борьба с последствиями снижения функции яичников, как с точки зрения количества, так и с точки зрения качества половых клеток, является основной задачей современных клиник по лечению бесплодия, и поэтому много научных усилий прилагается для оптимизации и расширения обоих этих аспектов. Однако в конечном счете достижения современной медицины в этом отношении были весьма ограничены, за исключением разработки синтетических стероидов полвека назад и последующих усовершенствований в области очистки и синтеза гонадотропинов. В то время как клинический процесс ЭКО во многих отношениях стал революционным, его подход к снижению количества и качества яйцеклеток по-прежнему заключается в попытке преодолеть их за счет увеличения количества или возможности использовать яйцеклетки от другой, более молодой женщины.
Однако существует множество интересных областей исследовательской деятельности, которые действительно затрагивают эти фундаментальные вопросы, в частности, углубленное понимание физиологических факторов, которые регулируют поддержание здоровья яйцеклеток в спящем фолликуле и во время развития фолликула, регулирование самой активации роста фолликула и контроль текущего развития фолликула в конкуренции с другими факторами. Важно также изучать растущие фолликулы и молекулярные механизмы, которые обеспечивают точность доставки эуплоидных гамет ожидающим сперматозоидам.
В этом кратком обзоре будут рассмотрены некоторые ключевые аспекты функции яичников, которые лежат в основе ключевого аспекта фертильности – развития полноценной яйцеклетки.
Источник: Anderson RA, Marston AL, Telfer EE. Oocyte development: it's all about quality. Reprod Biomed Online. 2025 Apr;50(4):104804. doi: 10.1016/j.rbmo.2025.104804. PMID: 40287201; PMCID: PMC7617848.
Хотя основное внимание как в исследованиях, так и в клинической практике вспомогательной репродукции уделяется эуплоидии яйцеклетки, одним из физиологически примечательных аспектов является то, что она возникает задолго до того, как требуется фертильность, на ранних стадиях внутриутробной жизни. После периода пролиферации половых клеток в конце первого триместра, вызванного не до конца понятыми сигналами, но включающими передачу сигналов ретиноевой кислотой и РНК-связывающие белки, оогонии начинают вступать в мейоз.
В отличие от яичника грызунов, где эта волна мейотических процессов распространяется в продольном направлении вдоль яичника, у человека она начинается в более центральных оогониях, а те, что расположены по периферии яичника, остаются митотическими; таким образом, примерно к 15 неделям развития первые примордиальные фолликулы уже заметны, но все еще окружены продолжающейся оогониальной пролиферации.
Во время этой первой профазы мейоза устанавливаются хромосомные переходы, необходимые для генетического разнообразия, и их необходимо стабильно поддерживать до тех пор, пока мейоз не возобновится во время овуляции. Это достигается за счет того, что сестринские хроматиды удерживаются вместе благодаря сцеплению, которое устанавливается по всей длине хромосом при их репликации.
Когезия опосредуется кольцевидным белковым комплексом, называемым когезином, который в настоящее время становится все более понятным на молекулярном и биохимическом уровне, о чем будет рассказано ниже.
Генетические мутации в генах, лежащие в основе ключевых компонентов когезина, были идентифицированы как у женщин, так и у мужчин с дефектами гаметогенеза, приводящими к преждевременной недостаточности яичников и азооспермии, соответственно. В дополнение к этим внутренним факторам, способствующим формированию большого количества потенциально дееспособных половых клеток, яичник также будет подвержен влиянию окружающей среды, таким как загрязнение воздуха, воды и пищевых продуктов химическими веществами, а также ожирение, что, следовательно, потенциально влияет на здоровье внуков беременной женщины.
Таким образом, после этой ранней критической активности популяция примордиальных фолликулов формируется еще до рождения. Хотя существование оогоний в послеродовом яичнике, которые могут впоследствии образовывать примордиальные фолликулы, остается предметом дискуссий у людей, это четко установлено у многих видов, включая некоторых млекопитающих. Потенциальная ценность возможности стимулировать реактивацию этих клеток в дальнейшей репродуктивной жизни очевидна, но это остается дискутабельной областью репродуктивной медицины.
Исследования яичников в детском возрасте, проведенные в 1970-х годах и даже ранее, выявили значительную активность фолликулов, а более поздние исследования выявили продолжающиеся изменения в яичниках в течение этого периода, которые, вероятно, влияют на качество яйцеклеток у взрослых.
Морфологически яичник в детском возрасте содержит много примордиальных фолликулов с аномальными признаками, и хотя они очень распространены у детей младшего возраста и составляют, возможно, треть всех фолликулов, они постепенно утрачиваются, так что эти признаки никогда не наблюдаются в яичниках взрослых женщин. С функциональной точки зрения давно известно, что при беременности у женщин - подростков анеуплоидия встречается чаще, чем у женщин в возрасте 20 лет, и недавно был пролит свет на молекулярную основу этого явления в связи с повышенной частотой в метафазе I нерасхождения, происходящего в яйцеклетках у женщин в возрасте до 20 лет. А повышенная анеуплоидия у пожилых женщин возникает в результате преждевременного разделения сестринских хроматид и обратной сегрегации.
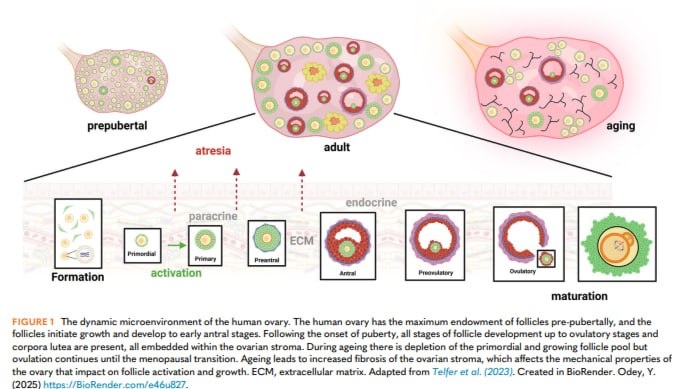
Также происходят прогрессирующие изменения в составе стромы яичников, которые начинаются в подростковом возрасте и прогрессируют в течение репродуктивной жизни до пострепродуктивного периода. Эти изменения, особенно в соотношении коллагена и эластина в составе стромы, приводят к повышению жесткости яичников в детском и подростковом возрасте и дальнейшему увеличению в более поздние репродуктивные годы. В настоящее время установлено, что это влияет на скорость активации роста фолликулов, при этом снижается активация примордиальных фолликулов в более жестких яичниках.
Важным открытием стало резкое увеличение фиброза яичников, связанное с возрастом, первоначально выявленное у мышей, но теперь подтвержденное также у женщин. Это также связано с усилением воспалительной реакции в яичниках, что является растущей областью исследований с пока неясными последствиями.
Однако на мышиной модели было продемонстрировано, что препараты, которые уменьшают фиброз и связанное с ним воспаление, независимо от того, связаны ли они с возрастом или даже с ожирением, могут повышать фертильность у пожилых животных, получавших лечение. Примечательно, что метформин активен в этом отношении, как и препараты, улучшающие функцию митохондрий. Это очень открытая область для исследований с потенциальной немедленной клинической пользой.
Примордиальные фолликулы являются фундаментальными структурами развития яичника млекопитающих, состоящими из ооцита, остановленного в профазе I мейоза, и соматических клеток (гранулёзы), остановленных на стадии G0 клеточного цикла, с взаимозависимостью на протяжении всего роста фолликула и созревания ооцита до момента овуляции. Примордиальные фолликулы могут (и большинству из них это необходимо) оставаться в состоянии покоя в течение многих лет, прежде чем активируются для роста и развития зрелого ооцита. Длительный покой представляет собой серьезную проблему для поддержания качества, поскольку ооциты необходимо защищать от таких стрессоров, как активные формы кислорода (АФК), которые являются побочными продуктами клеточного метаболизма, высокоактивными и вызывают окислительное повреждение компонентов клетки.
Недавние исследования показали, что у спящих ооцитов развился механизм ограничения повреждения АФК путем подавления митохондриального комплекса 1, ключевого генератора АФК в клетке. Отсутствие комплекса 1 приводит к менее эффективному клеточному метаболизму, но, поскольку этот метаболизм низок в примордиальных фолликулах, по-видимому, это стоит того, чтобы заплатить за преимущество снижения продукции АФК. Как только фолликулы активируются к росту, АФК становятся значительным фактором, влияющим на качество ооцитов, и снижение этого фактора стало ключевой терапевтической целью для смягчения последствий старения.
Дополнительные механизмы защиты примордиальных фолликулов были обнаружены после того, как было выявлено, что в яйцеклетках и соматических клетках примордиальных фолликулов мышей содержатся долгоживущие белки, которые уменьшают повреждения в зародышевой линии. Некоторые из белков, которые считаются долгоживущими, включают TWINKLE, митохондриальную ДНК - геликазу, необходимую для репликации митохондриальной ДНК, BOP1 (фактор биогенеза рибосом BOP1), необходимый для сборки рибосом, и родственные когезину белки.
Количество примордиальных фолликулов постепенно и непрерывно уменьшается с момента рождения при постоянной активации роста фолликулов. Переход от примордиальных фолликулов к первичным является ключевым этапом и характеризуется дифференцировкой и пролиферацией гранулезных клеток с образованием единого слоя клеток кубической формы, окружающих яйцеклетку. Во время последующего развития фолликула яйцеклетка также растет и приобретает необходимую ядерную и цитоплазматическую активность, чтобы поддержать возобновление мейоза и обеспечить возможность оплодотворения и раннего эмбрионального развития. Это зависит от межклеточной связи между растущей яйцеклеткой и окружающими ее соматическими (гранулезными) клетками. Динамические двунаправленные коммуникации формируются между ооцитами и компартментами соматических клеток посредством установления физических связей и секреции аутокринных и паракринных факторов роста. Эти связи необходимы для прохождения всех стадий развития, начиная с начала роста фолликулов. Образуются тесные межклеточные связи и трансзональные выступы – узкие цитоплазматические отростки, похожие на филоподии, пересекают прозрачную оболочку от клеток гранулезы до соединения с плазматической мембраной яйцеклетки с помощью щелевых соединений на их концах, облегчающих диффузию факторов между соседними клетками. Эта функциональная сеть щелевых соединений между яйцеклеткой и клетками гранулезы и, кроме того, между самими клетками гранулезы поддерживает фолликул в функционально интегрированном состоянии и в конечном итоге определяет качество яйцеклетки.
Среди идентифицированных факторов, перемещающихся между яйцеклеткой и клетками гранулезы, растущая яйцеклетка выделяет представителей семейства трансформирующих факторов роста бета (TGFb), включая GDF9 (фактор дифференцировки роста 9) и BMP15 (костный морфогенетический белок 15). Эти ооцитарные факторы способствуют пролиферации клеток гранулезы и запускают переход от первичной ко вторичной стадии развития фолликула, а также формирование самих трансзональных выступов. Эти факторы, секретируемые яйцеклетками, имеют важное значение для регуляции раннего развития фолликулов, а также модулируют ФСГ-зависимую цитодифференцировку клеток гранулезы, метаболизм и рост кумулюса на более поздних стадиях.
Важность факторов, секретируемых яйцеклетками, очевидна у многих видов животных, включая человека, у которого мутации в GDF9 и BMP15 способствуют преждевременной недостаточности яичников.
Хотя связь между соматическими клетками и ооцитом двунаправленная и взаимозависимая, интригующее недавнее исследование показало, что хронологический возраст среды соматических клеток влияет на развитие и здоровье ооцита.
Старые ооциты мыши подвергались воздействию молодой фолликулярной среды и восстановили функциональность и улучшили качество, что позволяет предположить, что возраст соматических клеток играет решающую роль в качестве ооцитов. Молодая фолликулярная среда восстанавливала здоровье митохондрий в старых ооцитах, снижая концентрацию активных форм кислорода (АФК) и окислительное повреждение, которые являются ключевыми факторами, способствующими старению ооцитов. Считается, что молодые гранулезные клетки более эффективны в формировании эффективной коммуникационной сети с ооцитом, и что это отвечает за эффект омоложения; поэтому необходимо рассмотреть цели по улучшению коммуникационной сети внутри фолликула. Возможно, они могут быть терапевтической целью, и терапия, направленная на соматические клетки, может иметь преимущества по сравнению с прямым воздействием на ооциты.
Как упоминалось выше, значимым фактором снижения качества ооцитов с возрастом является окислительный стресс. Влияние старения на молекулярные механизмы, регулирующие функцию ооцита, остается недостаточно изученным; однако исследования на животных показали, что с возрастом снижается уровень внутренних регуляторов функции митохондрий и окислительного стресса в яичниках, таких как представители семейства сиртуинов. Это снижение связано со снижением концентрации никотинамидадениндинуклеотида (НАД+), фермента, состоящего из аденина и никотинамида, который участвует в цепи переноса электронов в митохондриях и является важной сигнальной молекулой, способной влиять на клеточную транскрипцию белков и репарацию ДНК. Существует значительный интерес к этому потенциальному подходу к широкому спектру возрастных заболеваний. Добавление НАД+ через метаболический предшественник никотинамид мононуклеотида к старым мышам привело к улучшению качества яйцеклеток с уменьшением аномальных мейотических хромосомных веретен и увеличению количества рожденных детенышей. Кроме того, повышение концентрации НАД+ у мышей, получавших гонадотоксичные химиотерапевтические препараты, привело к улучшению функции яичников, что отразилось как на количестве ооцитов, так и на количестве детенышей.
Когда фолликулы на стадии мультиламинарного развития образуют заполненное жидкостью пространство, называемое антральной полостью, это приводит к физическому разделению двух функционально различных популяций гранулезных клеток: муральных и кумулюсных. Клетки муральной гранулезы сохраняют более соматическую функцию в эндокринной регуляции, синтезируя ингибины и эстрогены, в то время как клетки кумулюса выполняют более гаметогенную роль в регуляции созревания ооцитов посредством паракринной регуляции. В то же время, стероидогенные тека-клетки, управляемые ЛГ, дифференцируются из слоёв стромальноподобных клеток вокруг базальной мембраны фолликула.
Ооциты приобретают способность возобновлять мейоз на ранних стадиях развития фолликула, при этом полная мейотическая компетентность достигается к моменту формирования антрального отдела. После этого ооциты необходимо удерживать в состоянии мейотической остановки до тех пор, пока они не получат соответствующие сигналы созревания при овуляции. Это достигается за счёт поддержания повышенной концентрации циклического АМФ в ооците и наличия циклического ГМФ в гранулёзных клетках, а также зависит от поддержания хорошей коммуникации между ооцитом и соседними соматическими клетками. Возобновление мейоза запускается всплеском гипофизарного ЛГ в середине цикла, который воздействует на муральные гранулёзные клетки посредством высвобождения белков, связанных с эпидермальным фактором роста, которые затем также действуют как на кумулюс, так и на ооцит, влияя на созревание ядра, компетентность ооцита и овуляцию.
Постоянно увеличивающийся возраст деторождения вывел хромосомные ошибки ооцитов на передний план в практике репродуктивной медицины.
Даже в пиковом фертильном возрасте (20–32 года) около 20% человеческих ооцитов являются анеуплоидными, а после 32 лет их число увеличивается до более чем 50%, что приводит к параллельному увеличению анеуплоидии эмбрионов. В понимании механистической причины этого эффекта материнского возраста большое внимание было уделено когезину. Как описано выше, когезин захватывает две сестринские хроматиды во время репликации ДНК, которая в ооцитах млекопитающих происходит у плода. Когезин играет ключевую роль в обеспечении правильного распределения хромосом в ооците, и эксперименты на модельных организмах привели к пониманию того, как когезин поддерживает сегрегацию хромосом в оптимальных условиях. Во время профазы мейоза когезин поддерживает процесс мейотической рекомбинации, при которой хромосомы разрываются и склеиваются, что в конечном итоге приводит к образованию кроссинговеров. Кроссинговеры превращаются в хиазмы, которые являются физическими связями между гомологами. Важно отметить, что когезин на плечах хромосом стабилизирует хиазму, а когезиновое сцепление с хиазмой позволяет гомологичным хромосомам прикрепляться к веретену деления с противоположных полюсов, создавая натяжение, противодействующее силе микротрубочек. В конечном итоге, и в идеале, только когда все хромосомы правильно прикрепились таким образом, фермент сепараза активируется и разрезает когезин, запуская движение гомологов к противоположным полюсам. Однако когезин защищен от сепаразозависимого расщепления в перицентромерной области вокруг центромер белком шугошином (SGO2). Этот прицентромерный когезин важен для прикрепления сестринских хроматид к микротрубочкам с противоположных полюсов в мейозе II, и его последующая потеря при оплодотворении в результате второго раунда активации сепаразы приводит к равномерному расхождению сестринских хроматид к противоположным полюсам.
Следовательно, ухудшение работы когезина может привести к неправильному расхождению хромосом в мейозе I или II. На практике у людей наблюдаются ошибки в обоих мейотических делениях, но преждевременная потеря когезии сестринских хроматид преобладает при увеличении возраста матери.
Существенные доказательства, полученные на модельных организмах, указывают на то, что ухудшение когезина является потенциальным фактором, способствующим увеличению анеуплоидии в ооцитах женщин старшего возраста. Эксперименты на ооцитах мышей не выявили свидетельств оборота когезина после профазы мейоза.
Предполагается, что когезин, накопленный в плоде, должен удерживать хромосомы вместе на протяжении всей жизни особи до овуляции этого ооцита, что, возможно, происходит 40 или более лет спустя у человека. Однако недавно у плодовой мушки Drosophila было обнаружено образование когезина после S-фазы мейоза, что демонстрирует возможность восстановления когезина. Тем не менее, многие исследования показали, что уровень хромосомного когезина снижается с возрастом в ооцитах мыши и человека, и что расстояние между кинетохорами сестринских хроматид увеличивается с возрастом, что указывает на потерю когезии. Кроме того, частичное истощение во многих модельных системах приводит к анеуплоидии, напоминающей наблюдаемую при увеличении возраста матери, что подтверждает идею о том, что ухудшение состояния когезина является способствующим фактором. Кроме того, недавно обнаружено, что протектор когезина шугошин снижается в перицентромерах ооцитов человека, что привело к модели, согласно которой ослабление когезина приводит к снижению набора шугошина, что, в свою очередь, делает остаточный когезин уже уязвимым к расщеплению в мейозе I. Этот тип петли обратной связи может объяснить, почему увеличение анеуплоидии ооцитов с возрастом матери происходит не постепенно, а резко примерно после 35 лет.
Ключевые вопросы, которые ещё предстоит решить, включают в себя:
- Почему когезин особенно склонен к ухудшению?
- Является ли это следствием отсутствия обновления, или обновление происходит, но неэффективно?
- И есть ли потенциал для активации путей омоложения когезина для восстановления функциональности старых ооцитов и улучшения фертильности, как естественной, так и при помощи репродуктивных технологий?
Хотя ответы на эти вопросы должны быть приоритетными для дальнейшего изучения, не следует игнорировать возрастные эффекты на другие компоненты механизма сегрегации.
#развитие_яйцеклеток_япп