
Заражённость подсолнечника заразихой была впервые зарегистрирована в конце XIX века, когда началось массовое возделывание этой культуры. Паразит, мигрировавший на масличные растения из полыни, стал источником серьёзных проблем для сельхозпроизводителей. Начиная с середины XX века, подсолнечная заразиха, вызванная паразитическим растением Orobanche cumana, демонстрирует постоянную эволюцию, являясь реакцией на селекционные усилия по созданию устойчивых сортов подсолнечника. Исследования, проведённые на разных континентах, позволили выделить несколько рас этого паразита, каждая из которых обладает уникальным набором вирулентных свойств и датируется появлением в различное время.
При применении устойчивых сортов подсолнечника с определёнными R генами (генами устойчивости) паразит оказывается под сильным селективным давлением. Согласно модели «Ген на Ген», каждая пара
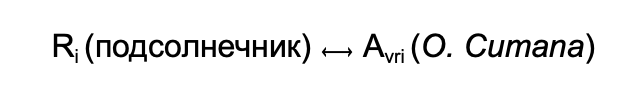
определяет результат заражения. Если в геноме паразита происходят мутации или рекомбинация, которые приводят к изменению или отсутствию Avr гена, возникает новая раса, способная обходить защиту растения.
Как показывает эволюция заразихи, механизмы вертикальной устойчивости подсолнечника легко преодолимы паразитом. Исследования показали, что популяции, паразитирующие на подсолнечнике, характеризуются низким внутрипопуляционным разнообразием и в целом низкой дифференциацией между популяциями. Кроме того, были предложены различные механизмы для объяснения эволюции заразихи, такие как мутации единственного гена в локальной популяциях или генетическая рекомбинация авирулентных генов.
Современные исследования, основанные на молекулярном анализе, подтверждают наличие последовательной эволюционной линии от расы «A» до «E» и новейших агрессивных и вирулентных рас «F», «G» и «Н». В некоторых источниках встречается подозрение на появление новых современных рас подсолнечниковой заразихи более агрессивных и вирулентных «I» и «J».
Эволюция рас заразихи
Таким образом, на данный момент выделяют и характеризуют следующие этапы эволюции рас O. Cumana:
- Раса А. Это «исходный» тип паразита, встречающийся на ранних этапах интенсивного выращивания подсолнечника, для которого характерно базовое взаимодействие с растением-хозяином. Является исходным типом, перешедшей на подсолнечник от полыни и впервые была зафиксирована в России в 1882 году. Первые сорта устойчивые к данной расе были выведены в России в начале ХХ века, данные сорта носили ген Or1.
- Раса В. Это адаптация к первым сортам подсолнечника с R-генами (Or1). Было обнаружено что что заразиха расы В неоднородна по составу и различна по вирулентности, раса состоит из 12 биотипных рас, которые были зафиксированные в 1925 году. Она способна идентифицировать те сорта, которые уже демонстрировали устойчивость к расе А. В 1934 году были выведены новые сорта с включённым геном устойчивости Or2.
- Раса С. Молдовский тип. Впервые была определена в 1963 году в Молдавской ССР. Данная раса возникла как ответ на дальнейшее применение устойчивых сортов, а затем она распространилась вдоль черноморского побережья. Ее вирулентность обусловлена изменениями в наборе Avri-генов, что позволило ей обходить защитные механизмы сортов подсолнечника с Or1 - Or2 генами. В СССР в 1976 году были получены сорта подсолнечника устойчивые к новой расе С.
- Раса D. Была обнаружена в конце 80-х годах ХХ века и имела усиленную вирулентность и отмечалась во многих странах почти одновременно. У данной расы наблюдается усиление агрессивных свойств, что приводит к быстрому заражению и значительным потерям урожая на полях, где ранее использовались сорта, устойчивые к расам A–C имевшие ген устойчивости Or1 - Or3.
- Раса Е. Как и раса D – морфологически подобна расе С, обнаруженная в середине 90-х годах ХХ века. Раса способна обходить новые комплексы устойчивости, сформированные на основе накопленного опыта борьбы с предыдущими расами. Проявляется при использовании сортов и гибридов подсолнечника с геном устойчивости Or1 – Or4.
- Раса F. Была идентифицирована в 1997 году в Румынии и Испании с высокой агрессивностью. Раса демонстрирует более высокую вирулентность и адаптивность, что свидетельствует о быстром темпе эволюционных изменений. Считается первой высоковирулентной расой. Она обходить защитные механизмы сортов и гибридов подсолнечника с Or1 – Or5 генами.
- Раса G. Достаточно агрессивная мутация локальных экземпляров заразихи, идентифицирована в середине 00-х ХХI века. Считается самой распространённой на данный момент. Поражает гибриды имеющие защитные гены Or1 — Or5, обеспечивающие устойчивость к расам O. cumana от A до E и ген HaOr7 (кодирует рецепторную протеинкиназу с повторами богатыми лейцином тем самым обеспечивает устойчивость, препятствуя прикреплению O. cumana к сосудистой системе корней) к расе F.
- Раса Н. Самая агрессивная локальная мутация заразихи. Идентифицирована в середине 00-х ХХI века вместе с расой G. Эти расы демонстрируют еще более высокую вирулентность и адаптивность, что свидетельствует о быстром темпе эволюционных изменений под воздействием однородных селекционных программ. Идентифицировать можно при возделывании гибридов имеющий большой механизм генной защиты Or1 - Or5, обеспечивающие устойчивость к расам от A до E и ген HaOr7 к расе F а также OrDeb2 и OrAnom1 к расе G.
- Раса I. Существует информация об идентификации данной расы, но пока она не имеет официального научного подтверждения.
Появление вирулентных рас вызвана развитием доминантных генов, поэтому каждый раз, когда селекционеры находят источник устойчивости, паразит образует неизвестный вирулентный вид.
Несколько лет назад компания BASF совместно с ВНИИМК провела серьезное исследование рас заразихи в 3-х основных регионах, высевающих подсолнечник в России - Черноземье, Юге России и Поволжье.
Из 26 точек отбора образцов в регионах Черноземье, на Юге России и Поволжье в 13 присутствовали образцы одной из самых вирулентных рас - H, это 50 %! Кроме того, было выявлено, что в полях присутствует не одна раса, а как правило смесь из 2-3 рас!
Сложности борьбы с заразихой
Основная сложность в борьбе с паразитом заключается в том, что мутации рас заразихи происходят на локальном уровне, 70% генетических различий происходит внутри рас, а только 30% – между ними.
Передача генетического материала между расами может влиять на частоты генов и помогать заразихе адаптироваться к своему хозяину, вводя новые полезные мутации. Это означает, что заразиха может преодолеть устойчивость семи расовому гибридов в одном регионе, но не сможет сделать это с другими гибридами в другом.
Расы одного региона отличаются от рас в других, но могут иметь как сходства, так и различия, образуя разные группы. Высокая изменчивость внутри рас объясняется тем, что растения из разных сельскохозяйственных регионов принадлежат к одной расе, но имеют разный генетический состав. Поэтому их нельзя классифицировать как одну, вторую или третью расу, так как они не являются генетически идентичными.
Цикл развития заразихи мы рассмотрим в следующем выпуске. Не пропустите!