Краткий очерк
Азендозавр, или же азендохзавр (лат. Azendohsaurus, «ящер из Азендо»), – вымерший род архозавроморфов аллокотозавров из семейства азендозаврид, живший на территории Марокко и Мадагаскара в среднем-позднем триасе: с ладинского по карнийский ярусы.

Азендозавр является типовым родом в семействе азендозаврид, названном в честь него.
Виды
Азендозавр известен по двум видам:
1. Azendohsaurus laaroussii, – типовой вид из Марокко;
2. Azendohsaurus madagaskarensis, вид собственно из Мадагаскара.
Почему неудавшийся?
Действительно, почему азендозавр неудавшийся динозавр, если он и динозавром, то собственно не был?
Тут, как и всегда, замешана штука под названием конвергенция, или же конвергентная эволюция, которая делает похожими разных животных, даже необязательно родственных друг другу.
Так, строение челюстей и зубов у азендозавра, которые имеют производные признаки, похожие на аналогичные у растительноядных динозавров. Так азендозавра записывали сначала в орнитоподы, а потом в прозауроподов, или же зауроподоморфов.
Впрочем, строение скелета азендозавра, который обнаружили чуть позже, демонстрировало признаки присущие базальным архозавроморфам, а следовательно динозавром он быть не мог.
Так почему же азендохзавр имел похожие черты с завроподами? А об этом чуть ниже.
Описание
Размеры
Азендозавр был достаточно крупным: 2-3 метра в длину.
Череп
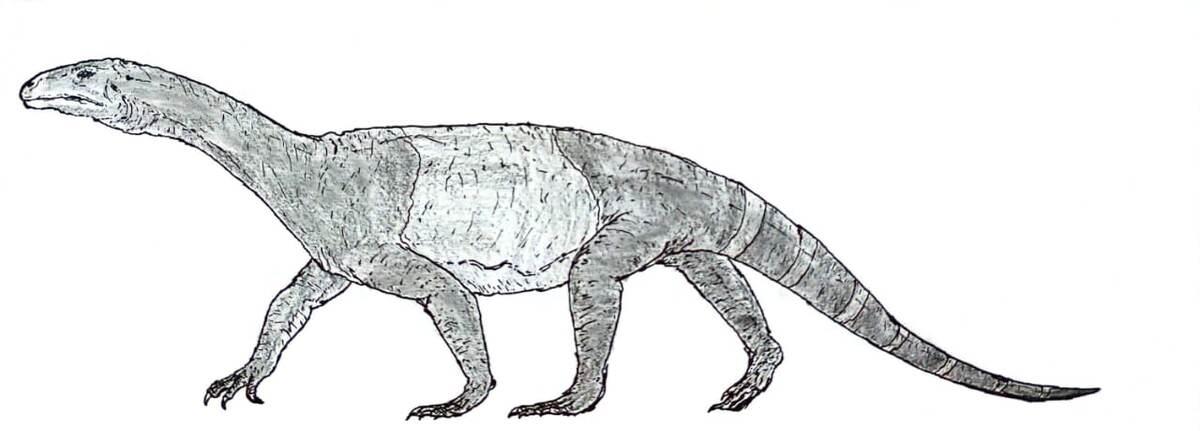
Череп азендозавра вида A.madagaskarensis известен почти полностью. Он имеет крепкое строение, короткую и квадратную форму и глубокую морду.Предчелюстные кости плавно изогнуты в передней части верхней челюсти, образуя тупой круглый кончик морды, а нижние челюсти имеют глубокий загнутый книзу кончик, как у зауроподов. Костные ноздри сливаются в одно (конфузное) отверстие, направленное вперёд, в передней части морды, как у ринхозавров.
Череп имеет ряд общих черт с зауроподоморфами, в том числе загнутую вниз нижнюю челюсть, мощный дорсальный отросток верхней челюсти и некоторые особенности зубов. Отросток на верхней челюсти обычно указывает на наличие анторбитального отверстия у архозавров, но у азендозавра это пространство занято слезной костью перед глазами. Это уникальное строение, не известное другим архозавроморфам триасового периода, за исключением родственного шрингазавра.Орбиты почти полностью заполнены крупными склеротическими кольцами, что указывает на большие глаза.Нижнее височное отверстие открыто снизу, разделяя скуловую и квадратно-скуловую кости (примитивный признак для архозавроморфов). Как и у других ранних архозавроморфов, у азендозавра есть небольшое (3–5 мм в диаметре) теменное отверстие («третий глаз») на крыше черепа.
Нижняя челюсть особенно похожа на челюсти зауроподоморфов: суставной шарнир, на котором крепится челюсть, расположен ниже уровня зубного ряда, а нижние челюсти изогнуты вниз, как и зубы аналогичной формы. Эти особенности встречаются у других травоядных архозавроморфов триасового периода, но такое сочетание известно только у азендозавра и зауроподоморфов.
Все зубы имеют примерно листовидную (ланцетовидную) форму с расширенными коронками и выпуклыми основаниями, которые срастаются с челюстными костями (анкилотекодонты).Однако верхние и нижние зубы явно гетеродонтные, и их легко отличить друг от друга. Верхние зубы относительно короткие и широкие у основания, с 4–6 бугорками на каждой поверхности, как у птицетазовых; нижние зубы почти в два раза выше и имеют в два раза больше бугорков, что больше напоминает зубы зауроподоморфов. Четыре предчелюстных зуба — самые длинные зубы в верхней челюсти, и они более загнуты назад, чем остальные.
Нёбо необычно покрыто многочисленными полностью сформировавшимися нёбными зубами, которых может быть до четырёх наборов на крыловидной кости и дополнительных рядов на нёбной кости и вомерах. У взрослых представителей A.madagaskarensis есть по меньшей мере 44 пары нёбных зубов в дополнение к 4 зубам в каждой предчелюстной кости и 11–13 зубам в каждой верхнечелюстной кости, а также максимум 17 зубов в зубной кости. Небные зубы не являются редкостью у травоядных рептилий, но у азендозавра они почти такой же формы, как и те, что расположены по краям челюсти, но немного толще. У других архозавроморфов с небными зубами небный зубной ряд состоит из более простых мелких куполообразных зубов.
Единственным описанным материалом азендозавра A.laaroussii являются зубные кости, верхнечелюстные кости, предчелюстная кость и несколько зубов. По общей форме они напоминают A. madagaskarensis, но имеют несколько отличительных особенностей. У A. laaroussii больше зубов: 15–16 на верхнечелюстной кости по сравнению с 11–13 у A. madagaskarensis. Зубы A. laaroussii также выше, чем у A. madagaskarensis, и имеют более плотно расположенные зубчики. Ещё одним отличием этих двух видов является наличие заметного киля на внутренней поверхности верхней челюсти. У A. laaroussii этот киль проходит по всей длине верхней челюсти, а у A. madagaskarensis — только по задней её части. Любые другие возможные различия между этими двумя видами невозможно определить без останков черепа и скелета.
Скелет
Большая часть позвоночника азендозавра известна, и, хотя он сохранился не полностью, предполагается, что у него было 24 предкрестцовых позвонка (включая атлант и ось). Крестец в области бёдер состоит всего из двух позвонков, а полное количество хвостовых позвонков в хвосте неизвестно, но, по оценкам, их всего около 45–55 (мало для архозавра).
Шейные позвонки меняют форму по мере продвижения вниз по шее, начиная с характерного удлинения с длинными и низкими остистыми отростками и постепенно укорачиваясь к основанию шеи, но при этом становясь всё более высокими и узкими. Такое укорочение наблюдается в шеях других аллокотозавров, таких как трилофозавр, но не встречается у других длинношеих архозавроморфов (например, у танистрофеид средние шейные позвонки самые длинные). Шея была приподнята над телом, о чём свидетельствует наклонный угол зигапофизов, соединяющих каждый позвонок, а также то, что передние зигапофизы каждого позвонка расположены выше, чем задние. Шея, вероятно, была изогнута дугой, что обусловлено строением шейных позвонков в этом положении.
Грудные позвонки спины в целом похожи на последние шейные позвонки, у них высокие вертикальные остистые отростки. Длина этих позвонков также уменьшается по направлению к спине, но не так резко, как в случае с шеей. Однако последний грудной позвонок уникален тем, что его остистый отросток направлен вперёд. Из двух крестцовых позвонков первый крупнее и прочнее, с высокими остистыми отростками в задней части. Оба крестцовых позвонка имеют крупные рёбра, полностью сросшиеся с позвонками, которые соединяются с подвздошными костями.
Хвостовые позвонки похожи на другие позвонки, но с наклонёнными назад остистыми отростками. Длина хвостовых позвонков и высота остистых отростков постепенно уменьшаются по направлению к кончику хвоста, в отличие от некоторых других архозавроморфов, у которых позвонки удлиняются к концу. Это означает, что хвост был коротким и не сужался к концу, но сам кончик хвоста неизвестен. Начиная с 3-го или 4-го позвонка и до последнего известного хвостового позвонка в ряду у них неизменно присутствуют шевроны.
Шейные рёбра длинные и тонкие, становятся более крепкими и сужаются по мере продвижения вниз по шее. Некоторые шейные рёбра в нижней половине шеи имеют на внутренней поверхности небольшую фасетку на кончике, которая могла удерживать кончик предыдущего ребра, образуя жёсткий ряд шейных рёбер (что также предполагается для длинношеего танистрофея), который придавал шее жёсткость.Грудные рёбра длинные и изогнуты наружу, что указывает на то, что у азендозавра была широкая и глубокая грудная клетка бочкообразной формы. Длина и кривизна рёбер уменьшаются по направлению к хвосту, а последнее ребро короткое, полностью сросшееся с последним спинным позвонком и направленное прямо в стороны. Для азендозавра известен только один набор гастролитов, и их очень хрупкое строение и редкость по сравнению с другими костями позволяют предположить, что у него не было хорошо развитой корзины гастролитов под брюхом.
Конечности и пояса
Передние конечности и плечи (грудной пояс) азендозавра хорошо развиты и крепки.Лопатка (плечевая кость) длинная, примерно в два раза выше, чем шире, и соответствует длине и изгибу рёбер, что позволяет ей вмещать глубокую грудную клетку. Лопатка вогнута с обеих сторон, а её слегка расширенный кончик направлен назад. Межключичная кость крупная и прочная, как у трилофозавра и некоторых ринхозавров, с длинным «лопатообразным» задним отростком, который уплощён и расширен к концу. У неё также есть уникальный направленный вперёд отросток, который есть только у проторозавра и некоторых ранних диапсид (у большинства других архозавроморфов вместо него выемка).
Клювовидные кости крупные и округлые, соединяются с лопаткой, образуя суставную впадину (плечевой сустав). Суставная впадина обращена вбок, что характерно для рептилий, лежащих на земле, однако лопаточная часть направлена немного назад, что может указывать на то, что плечевая кость находилась в более приподнятом положении. Сама плечевая кость крупная и широкая с обоих концов, с относительно узкой «талией» в середине и очень хорошо развитым дельтопекторальным гребнем. Лучевая кость такая же массивная, с немного расширенными концами, в то время как локтевая кость сильно расширена с обоих концов, хотя и в меньшей степени дистально.
Бёдра (тазовый пояс) не такие широкие, как плечи, а три тазовые кости примерно одинакового размера.Подвздошная кость высокая и изогнутая по верхней поверхности, с коротким округлым отростком спереди и более длинным заострённым отростком сзади.Лобок направлен вниз и немного вперёд, а на его конце есть лишь небольшое утолщение (бугорок).Седалищная кость относительно короткая, короче подвздошной кости, и имеет примерно треугольную форму с прямыми краями и закруглённым задним концом. Суставные поверхности между каждой седалищной костью необычно расширены по сравнению с другими архозавроморфами. Все три кости участвуют в формировании глубокой округлой вертлужной впадины (тазобедренного сустава). В отличие от открытого сустава у динозавров, внутренняя стенка вертлужной впадины у азендозавра представляет собой цельную кость.
Большие крестцовые рёбра соединяются с подвздошной костью таким образом, что она располагается почти вертикально, хотя из-за небольшого угла наклона вниз вертлужная впадина была направлена не только в сторону от тела, но и вниз примерно на 10–25° от вертикали.Бедренная кость длинная и имеет S-образную форму, с немного расширенной головкой, которая не повёрнута внутрь, в отличие от костей динозавров, что указывает на то, что она не располагалась вертикально. Бедренная кость также скручена вдоль своей оси, так что поверхности головки и колена смещены относительно друг друга примерно на ~75°.Большеберцовая кость составляет примерно 75 % длины бедренной кости, слегка изогнута и очень прочная по сравнению с костями других архозавроморфов, за исключением самых крупных ринхозавров.Малоберцовая кость, напротив, тонкая и сильнее скручена вдоль своей оси.
Конечности азендозавра хорошо сохранились в окаменелостях, включая полную кисть (manus) и стопу (pes) в сочленении. Все кости запястья и кости предплюсны хорошо окостенели и чётко различимы, а сложная предплюсна состоит из девяти костей. Пястные кости в кисти примечательны тем, что они расходятся по плавной дуге, а длина пальцев почти симметрична относительно длинного третьего пальца, а также относительно нерасходящихся первого и пятого пальцев. Это отличает их от кистей других рептилий, у которых первый и пятый пальцы расположены далеко друг от друга, а четвёртый палец самый длинный. Плюсневые кости и пальцы стопы также расходятся по плавной дуге, но, в отличие от кисти, они несимметричны: четвёртый палец длинный, а пятый — короткий и загнутый.
Все пальцы на руках и ногах необычайно короткие для архозавроморфов, в отличие от родственного вида трилофозавра.Когти (или подушечки) очень большие, узкие и резко загнутые, они значительно крупнее предыдущей фаланги, к которой были прикреплены.Пальцы и когти имеют общие черты с пальцами и когтями дромеозаврид и троодонтид манирапторов, а также других рептилий, таких как черепаха проганохелис. Эти общие черты связаны с хорошо развитыми сухожилиями сгибателей, и предполагается, что это адаптация для противостояния силам, возникающим при рытье.
Образ жизни
Что ел и как ел
По вышенаписанному можно догадаться что азендозавр был животным растительноядным, поэтому и приспособления для поедания растений у него были. В случае азендозавра таковыми были его листовидные зубы, которые, судя по микроповреждениям (следам оставленным на поверхности зубов во время питания), были предназначены для поедания мягкой растительности. Характер микроизноса также указывает на то, что для пережёвывания пищи он использовал простые движения челюстью вверх и вниз.
Полностью сформировавшиеся нёбные зубы позволяют предположить, что они использовались для специализированного питания. Однако функциональные исследования нёбных зубов не проводились, поэтому точно неизвестно, для чего они использовались, хотя их форма, схожая с формой краевых зубов, позволяет предположить, что они использовались для обработки похожей пищи. Крыловидная кость молодой особи A. madagaskarensis имеет меньшее количество рядов нёбных зубов, которые меньше по размеру, чем у более крупных взрослых особей. Это указывает на то, что азендозавр увеличивал количество и размер нёбных зубов по мере взросления. У молодых особей также было меньше зубных зубов, чем у взрослых, хотя разница была гораздо менее существенной по сравнению с нёбными зубами (16 против 17 у взрослых особей).
Положение тела
Поза тела, характерная для азендозавра, представляет собой нечто среднее между распластанной и полураспластанной. Задние конечности полностью вытянуты в стороны от тела, бедренная кость выпрямлена, а голень согнута под ней в колене на 90°, как у ящерицы. Однако передние конечности и плечевой пояс указывают на то, что передняя часть тела располагалась более вертикально, чем задняя, с частично направленной вниз плечевой впадиной и плечевой костью, которая больше подходила для того, чтобы держать её частично вертикально, и по форме напоминала кости зауроподоморфов. Это необычное сочетание говорит о том, что азендозавр стоял, приподняв переднюю часть тела над землёй, что в сочетании с его длинной изогнутой шеей и маленькой головой позволяло ему щипать траву, находясь относительно высоко над землёй, в отличие от современных ринхозавров и цинодонтов, которые щипали траву низко над землёй. Адаптация к высокому росту, возможно, объясняет сходство между азендозавром и зауроподоморфами, которые приобрели схожие черты в строении шеи, передних конечностей и позвоночника, чтобы занимать схожие ниши. Однако более приземистая поза азендохзавра, вероятно, не позволяла ему высоко поднимать голову, как это делали полностью прямоходящие зауроподоморфы.
Как жил
Азендозавр вида A.madagaskarensis был возможно, самым крупным животным в экосистеме формации Макай. Он занимал нишу крупноразмерного растительноядного, питавшегося листьями деревьев, что подтверждается его высоким положением тела, а так же его длинной шеей.
Другими растительнодяными в этой местности были траверсодонтиды: менадон и дададон, каннемейериобразные дицинодонты, представленные одним неописанным видом, и ринхозавр исалоринх. Главным хищником по видимому был пробайногнат чиникводон (C.kalanoro), который раньше считался чиникводонтисом.
Помимо этого тут было достаточно количество рептилий сфенодонтов, парарептилий проколофонид, неописанных динозавроморфов, миниатюрный лагерпетид конгонафон и один "загадочный архозавр" неопределённой классификации.
Метаболизм и рост
Гистологическое исследование микроскопической структуры костей 2019 года для которого использовались тонкие срезы из плечевой, бедренной и большеберцовой кости азендозавра было сделано для определения скорости роста этого архозавроморфа. В ходе исследования выяснилось, что плотность сосудов азендозавра сопоставима с таковыми у птиц и млекопитающих, а выявленные типы костной ткани — в частности, энергозатратная фиброзно-ламеллярная костная ткань — указывают на высокую скорость метаболизма в состоянии покоя, которая находится в диапазоне показателей современных птиц и млекопитающих. Это привело к выводу, то азендозавр был эндотермным ("теплокровным") животным. Высокая скорость метаболизма в состоянии покоя, аналогичная той, что была у азендозавра, была выявлена у других, более развитых архозавроморфов (таких как пролацерта), и анализ показал, что эндотермия могла быть присуща архозавроморфам ещё на этапе их общего предка с аллокотозаврами.
Палеопаталогия
Несмотря на множество образцов, которые были исследованы, у A. madagaskarensis была зарегистрирована только одна патология. Образец UA 7-16-99-620, один из трех сохранившихся межключичных, был деформирован так, что длинный задний отросток был резко согнут вправо, по сравнению с нормальными прямыми задними отростками двух других межключичных.