Появление первых нейронов у эмбриона — это сложный и строго регулируемый процесс, который происходит в ходе нейрогенеза.
Этот процесс начинается на ранних стадиях развития нервной системы и включает последовательность молекулярных и генетических событий.
Основные этапы появления первых нейронов:
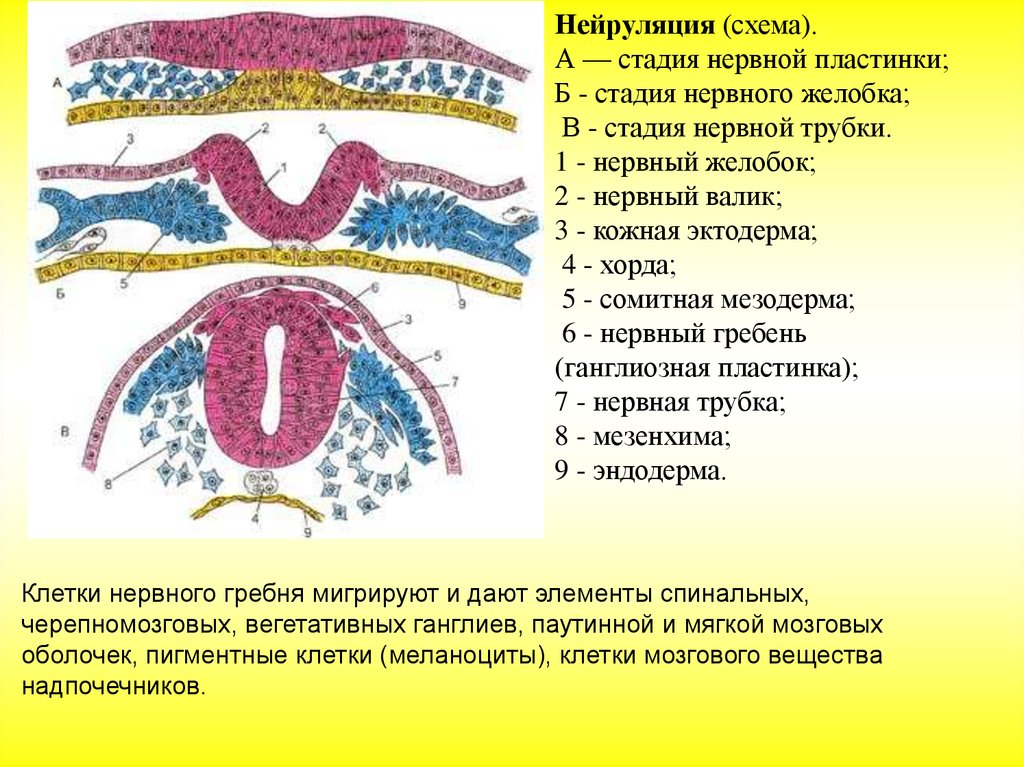
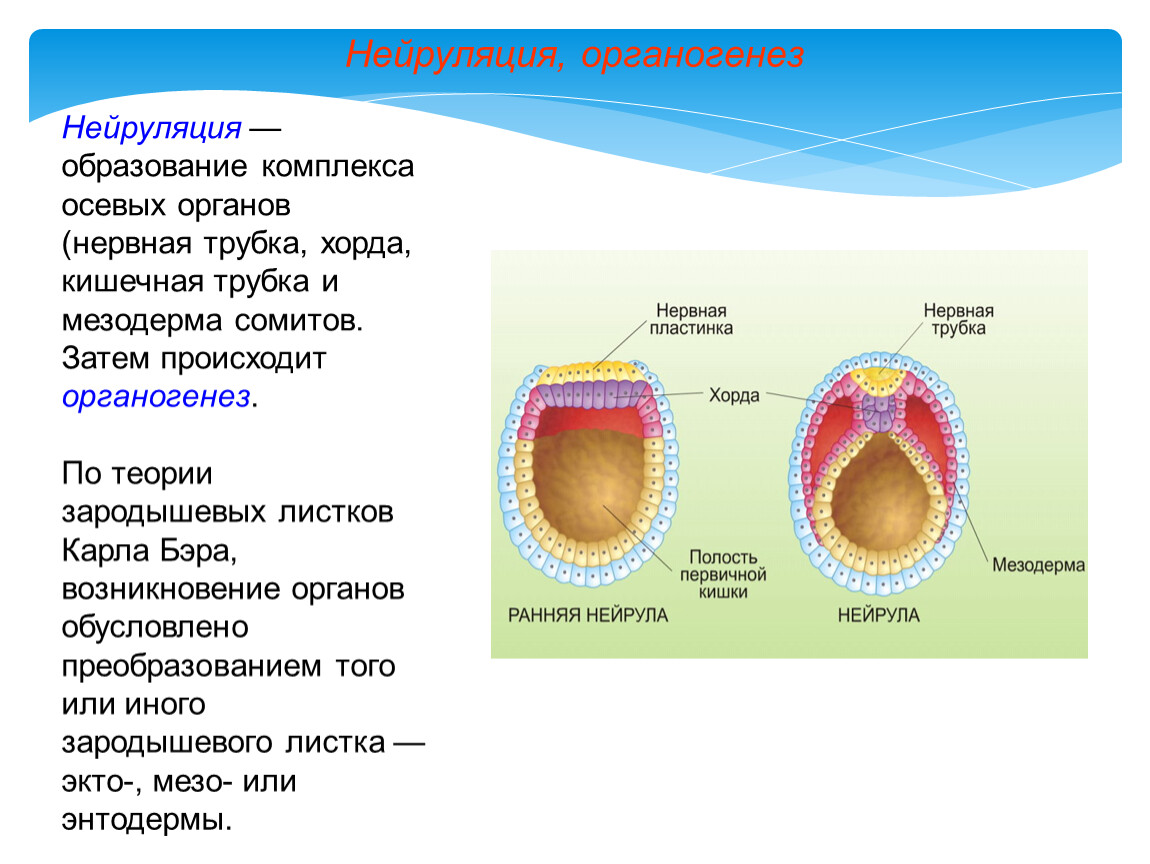
Нейруляция и образование нервной трубки
В начале эмбриогенеза формируется нервная пластинка, которая затем сворачивается в нервную трубку — зачаток центральной нервной системы. Клетки нервной трубки — это нейроэпителиальные клетки, которые являются предшественниками нейронов и глиальных клеток.
Нейруляция — процесс образования нервной трубки, предшественника головного и спинного мозга, в эмбриональном развитии позвоночных.
ai.mitup.ru
Процесс нейруляции включает несколько стадий:
- Образование на спинной стороне тела из клеток эктодермы плоской нервной пластинки. Этот процесс регулируется сигнальными молекулами, которые выделяют хорда и мезодерма.
- Формирование нервных валиков по краям нервной пластинки и нервной бороздки в центре.
- Смыкание нервных валиков и образование нервной трубки с полостью
( невроцелем) внутри. - Образование нервного гребня клетками, которые лежат между нервной пластинкой и кожной эктодермой.
- Миграция клеток нервного гребня. Поверхностный поток преобразуется в пигментные клетки кожи, внутренний — в чувствительные спинномозговые ганглии, симпатические нервные узлы, мозговое вещество надпочечников, парасимпатические ганглии.
Существуют разные типы нейруляции, которые различаются механизмами формирования нервной трубки:
- Первичная нейруляция — наиболее распространённый тип, характерен для большинства позвоночных. Нервная трубка образуется путём сворачивания и замыкания нервной пластинки.
- Вторичная нейруляция — наблюдается у некоторых позвоночных, в частности, в каудальной части тела. При вторичной нейруляции нервная трубка образуется путём конденсации мезенхимальных клеток с последующим образованием просвета.
На нейруляцию влияют генетические факторы и факторы окружающей среды, например дефицит фолиевой кислоты, воздействие тератогенов и инфекции
yandex.ru/search?text=Нейруляция+и+образование+нервной+трубки
Нейральное стволовое состояние
Нейроэпителиальные клетки сначала делятся пролиферативно, поддерживая популяцию стволовых клеток. Они экспрессируют гены, связанные с поддержанием стволового состояния, например, SOX2, PAX6, Nestin.
Нейрональная дифференцировка
Под влиянием внешних сигналов часть нейроэпителиальных клеток начинает выходить из цикла деления и дифференцироваться в нейроны.
Молекулярные и генетические механизмы
Внешние сигналы (морфогены)
BMP (Bone Morphogenetic Protein) и Wnt: при высоких концентрациях поддерживают эпителиальный стволовой статус. Их снижение в нервной пластинке способствует нейрогенезу.
Shh (Sonic Hedgehog): секретируется вентральными структурами нервной трубки и влияет на паттернизацию и дифференцировку нейронов.
FGF (Fibroblast Growth Factors): участвуют в поддержании нейрального стволового состояния и позже в индукции нейрональной дифференцировки.
Внутриклеточные транскрипционные факторы
Neurogenin (Neurog1, Neurog2) и Mash1 (Ascl1) — ключевые факторы, запускающие нейрональную программу дифференцировки. Они активируют экспрессию генов, ответственных за нейрональный фенотип.
Notch-сигналинг: поддерживает клетки в стволовом состоянии. При снижении активности Notch происходит активация нейрональных факторов.
REST (RE1-silencing transcription factor): подавляет нейрональные гены в стволовых клетках, при снижении REST начинается экспрессия нейрональных генов.
Генетические изменения
Активация нейрональных генов, таких как Tubb3 ( βIII-тубулин), Map2, NeuN — маркеры дифференцирующихся нейронов.
Изменения в эпигенетическом статусе ( метилирование ДНК, модификации гистонов) способствуют активации нейрональных генов и подавлению генов стволового состояния.
Первые нейроны появляются в результате комплексного взаимодействия внешних сигналов ( морфогенов),
внутриклеточных сигнальных путей и транскрипционных факторов, которые переключают нейроэпителиальные клетки из состояния пролиферации в дифференцировку.
На молекулярном уровне это сопровождается активацией нейрональных генов и подавлением генов, поддерживающих стволовой статус.
изучим возможные сигнальные пути или транскрипционные факторы
Расскажем подробнее о механизмах, которые обеспечивают переход нейроэпителиальных клеток к нейрональной дифференцировке, а также о последующих этапах формирования первых нейронов.
Дополнительные механизмы регуляции нейрогенеза
Роль Notch-сигналинга в поддержании баланса между пролиферацией и дифференцировкой
Notch-сигналинг активируется при взаимодействии рецептора Notch с лигандом (например, Delta или Jagged) на соседних клетках.
Активация Notch приводит к высвобождению внутриклеточного домена Notch (NICD), который перемещается в ядро и активирует гены семейства Hes (например, Hes1, Hes5).
Гены Hes подавляют транскрипцию нейрогенных факторов (Neurog1/2, Ascl1), поддерживая клетки в стволовом состоянии.
Снижение Notch-сигналинга (например, за счет уменьшения лигандов или обратной связи) приводит к снижению Hes и позволяет транскрипционным факторам нейрогенеза активировать нейрональные программы.
Транскрипционные факторы и их взаимодействия
Neurog2 и Ascl1 являются так называемыми "просоциализационными" факторами, которые активируют экспрессию генов, необходимых для формирования нейронального фенотипа: гены цитоскелета, синаптических белков, ионических каналов.
Эти факторы также могут усиливать экспрессию друг друга и подавлять гены, поддерживающие стволовое состояние.
Кроме того, они взаимодействуют с эпигенетическими модификаторами, чтобы изменить структуру хроматина, делая нейрональные гены более доступными для транскрипции.
Эпигенетические изменения
В процессе дифференцировки происходит деметилирование промоторов нейрональных генов и модификация гистонов (например, ацетилирование гистонов H3 и H4), что способствует открытию хроматина.
Активируются хроматин-модифицирующие ферменты, такие как гистондеацетилазы (HDAC), метилтрансферазы и др., которые регулируют доступ транскрипционных факторов к ДНК.
REST-комплекс, который подавляет нейрональные гены в стволовых клетках, сходит на нет, что позволяет экспрессии нейрональных генов.
Формирование первых нейронов
После активации нейрональных генов нейроэпителиальные клетки начинают выходить из клеточного цикла (G0-фаза) и начинают морфологическую дифференцировку: формируется аксоны, дендриты, происходит организация цитоскелета.
Экспрессия таких белков, как βIII-тубулин (Tubb3) и MAP2, способствует формированию микротрубочек и стабилизации нейрональных отростков.
Появляются первые синаптические белки, что позволяет нейронам устанавливать контакты друг с другом.
Эти ранние нейроны мигрируют к своим целевым областям в развивающемся мозге, где они продолжают созревать и формировать функциональные нейронные цепи.
Итог
Появление первых нейронов — результат тонко сбалансированного взаимодействия сигналов, транскрипционных факторов и эпигенетических механизмов, которые переключают нейроэпителиальные клетки от пролиферации к дифференцировке. Этот процесс обеспечивает формирование функциональных нейронов, необходимых для дальнейшего развития нервной системы.
Вот примеры конкретных генов и белков, участвующих в нейрогенезе, а также методы, которыми изучают эти процессы.
Примеры конкретных генов и белков
Гены и белки, поддерживающие стволовое состояние нейроэпителиальных клеток
SOX2 - транскрипционный фактор, поддерживает плюрипотентность и предотвращает преждевременную дифференцировку.
PAX6 - регулирует развитие нервной трубки и поддерживает нейроэпителиальные клетки.
Nestin - промежуточный филамент, маркер нейроэпителиальных стволовых клеток.
Hes1, Hes5 - эффекторные гены Notch-сигналинга, подавляют нейрогенные факторы.
Гены и белки, запускающие нейрональную дифференцировку
Neurog1, Neurog2 ( Neurogenins) - транскрипционные факторы, активирующие нейрональные гены.
Ascl1 ( Mash1) - транскрипционный фактор, способствующий нейрогенезу.
Tubb3 ( βIII-тубулин ) — структурный белок микротрубочек, маркер ранних нейронов.
MAP2 - белок, стабилизирующий микротрубочки в дендритах.
NeuN ( Rbfox3 ) — ядерный белок, маркер зрелых нейронов.
Эпигенетические регуляторы
REST ( RE1-silencing transcription factor) - репрессор нейрональных генов в стволовых клетках.
HDACs ( гистондеацетилазы) - ферменты, регулирующие ацетилирование гистонов.
DNMTs ( ДНК-метилтрансферазы) - ферменты метилирования ДНК, регулирующие экспрессию генов.
Экспериментальные методы изучения нейрогенеза
Молекулярно-биологические методы
RT-PCR и qPCR - измерение уровня транскриптов генов ( например - Neurog2, Ascl1, Hes1) для оценки их экспрессии на разных стадиях.
Вестерн- блоттинг - определение белков ( SOX2, βIII-тубулин) в клетках или тканях.
Иммуногистохимия и иммуноцитохимия - визуализация локализации белков с помощью антител (например, маркировка Nestin, NeuN в срезах мозга).
Генетические методы
Кокауты и нокауты генов ( CRISPR/Cas9, трансгенные модели ) - удаление или мутация генов (Neurog2, Hes1) для изучения их роли в нейрогенезе.
Линейное прослеживание клеток ( lineage tracing) - маркировка стволовых клеток и отслеживание их потомков,
чтобы понять, когда и как они дифференцируются.
Клеточные нейроструктуры
Нейросферы и нейрональные культуры из эмбриональных стволовых клеток -
in vitro модели для изучения этапов дифференцировки и влияния факторов роста (FGF, Shh).
Эксперименты с ингибиторами/активаторами сигналинговых путей - например,
блокирование Notch с помощью γ-секретазных ингибиторов для изучения влияния на нейрогенез.
Функциональные методы
Электрофизиология - регистрация активности первых нейронов для оценки их функциональной зрелости.
Конфокальная и двух-фотонная микроскопия - визуализация морфологии нейронов и синапсов в живых или фиксированных образцах.
Комбинируя эти методы, учёные могут детально изучать, как конкретные гены и белки регулируют переход от нейроэпителиальных клеток к первым нейронам, а также как эпигенетические механизмы и внешние сигналы влияют на этот процесс.
Это позволяет понять молекулярные основы развития нервной системы и искать пути лечения нейродегенеративных заболеваний.
Репрессор нейрональных генов — это белок, который подавляет экспрессию генов, необходимых для нейрональной дифференцировки, поддерживая клетки в недифференцированном состоянии.
Один из ключевых таких репрессоров — REST ( RE1-Silencing Transcription factor), также известный как NRSF ( Neuron-Restrictive Silencer Factor).
REST - основной репрессор нейрональных генов
Функция:
REST связывается с определёнными ДНК-последовательностями, называемыми RE1 ( Repressor Element 1),
расположенными в промоторах многих нейрональных генов.
При связывании REST привлекает комплекс ко-репрессоров и эпигенетических модификаторов (гистондеацетилазы - HDAC, метилтрансферазы ), что приводит к конденсации хроматина и подавлению транскрипции этих генов.
Роль в развитии:
В стволовых и предшественниках нейронов REST активно подавляет нейрональные гены, препятствуя преждевременной дифференцировке. По мере запуска нейрогенеза уровень REST снижается, что снимает репрессию и позволяет нейрональным генам (например, Tubb3, Syn1, Bdnf) экспрессироваться.
Регуляция REST:
Экспрессия REST и его активность регулируются на нескольких уровнях — транскрипционном, посттранскрипционном (например, микроРНК), а также через протеолиз и модификации белка.
Экспериментальное изучение REST
Измерение экспрессии REST:
RT-qPCR и вестерн-блоттинг позволяют определить уровень REST в разных клеточных популяциях (стволовые клетки, нейроны).
Хроматин-иммунопреципитация (ChIP):
используется для выявления связывания REST с промоторами нейрональных генов.
Кокауты и нокауты REST:
С помощью CRISPR/Cas9 или РНК-интерференции изучают эффект удаления REST - обычно приводит к преждевременной активации нейрональных программ и ускоренному нейрогенезу.
Использование ингибиторов HDAC:
Поскольку REST действует через ко-репрессоры, ингибиторы HDAC могут частично снимать репрессию REST.
REST - ключевой фактор, который предотвращает преждевременную активацию нейрональных генов в нерешённых клетках нервной системы.
Его регуляция позволяет точно контролировать время и место нейрогенеза, обеспечивая правильное развитие мозга.
Если хочешь, могу рассказать подробнее про механизмы взаимодействия REST с другими белками или про роль REST при заболеваниях нервной системы.
Ингибиторы — это вещества, которые уменьшают или полностью блокируют активность определённых белков или ферментов. В контексте регуляции нейрональных генов и нейрогенеза ингибиторы часто применяются для модуляции активности репрессоров и связанных с ними ко-репрессоров.
Основные функции ингибиторов в изучении репрессоров нейрональных генов
Ингибиторы гистондеацетилаз ( HDAC-инhibitors)
Механизм действия:
HDAC — ферменты, которые удаляют ацетильные группы с гистонов, вызывая конденсацию хроматина и подавление транскрипции.
Ингибиторы HDAC препятствуют этому процессу, способствуя ацетилированию гистонов, разрыхлению хроматина и активации транскрипции.
Роль в нейрогенезе:
Поскольку REST привлекает HDAC для подавления нейрональных генов, ингибиторы HDAC могут частично снимать репрессию REST, способствуя экспрессии нейрональных генов и стимулируя дифференцировку нейронов.
Примеры:
TSA ( триоксазол), валпроевая кислота, SAHA ( субероиланаксамид).
Ингибиторы протеаз
Используются для предотвращения деградации белков, включая REST или его ко-репрессоры, что помогает изучать стабильность и функции этих белков.
Ингибиторы сигналинговых путей
Например, ингибиторы Notch-сигналинга (γ-секретазные ингибиторы) могут косвенно влиять на экспрессию репрессоров или их мишеней, изменяя баланс между стволовыми и дифференцированными клетками.
Применение ингибиторов в исследованиях
Снятие репрессии:
Ингибиторы HDAC позволяют «разблокировать» нейрональные гены, что помогает понять роль эпигенетической регуляции в нейрогенезе.
Моделирование заболеваний: Некоторые нейродегенеративные заболевания связаны с нарушением регуляции генов; ингибиторы могут использоваться для коррекции экспрессии.
Терапевтический потенциал: HDAC-инhibitors исследуются как возможные препараты для стимуляции нейрогенеза и восстановления функций мозга.