Предпосылки открытия
В 1965 году было обнаружено, что первым продуктом фотосинтеза у сахарного тростника является не фосфоглицериновая кислота (кислота, имеющая углеродный скелет из 3х атомов углерода, т.е. С3-кислота), как у большинства растений умеренного климата, а С4-кислота. С тех пор выявили множество растений, в основном тропического и субтропического климата, у которых фотосинтез протекает по нестандартному пути. Такие растения получили название С4-растения. Примерами таких растений могут служить кукуруза, сахарный тростник, сорго и просо. Соответственно растения, в которых первым продуктом фотосинтеза является трёхуглеродная фосфоглицериновая кислоты, носят название С3-растений. Именно биохимию С3-растений рассматриваю при описании классического фотосинтеза. Но что же касается С4-фотосинтеза, именно об этом и будет эта статья.
История открытия
В 1966г. Был расшифрован генетический код, но помимо это события двое австралийских исследователя Хэтч и Слэк (Hatch, Slack) показали что С4-растения значительно эффективнее, чем обычные растения. Ученым стало интересно с чем это связано и они описали целый новый путь метаболизма углерода у С4-растений, получивший название путь Хэтча-Слэка.
Суть С4-фотосинтеза
С помощь особого белка (ФЕП) углекислый газ захватывается из атмосферы клетками мезофилла и под действием ферментов образуется С4-кислота (яблочная/щавелевая/аспарагиновая), затем образуется соль этой кислоты, которая поступает в особые клетки, окружающие проводящий пучок листа, так называемые клетки обкладки проводящего пучка, в хлоропластах которых соль С4-кислоты преращается в С3-кислоту, которая идёт на регенерацию изначального белка (ФЕП). При этом происходит увеличение концентрации углекислого газа в хлоропластах клетки обкладки проводящего пучка, который фиксируется уже обычным белком С3-фотосинтеза (РиБФ) и идёт уже по стандартному для фотосинтеза пути.
Примечание
Дорогой читатель, далее будет подробно расписан весь этот сложный процесс специально для тех, кто хочет погрузиться в него намного глубже. Если же, изложенного ранее материала предостаточно вам, то вы смело можете листать в конец статьи к абзацу с особенностями и заключению.
Более подробный механизм С4-фотосинтеза или путь Хетча-Слэка
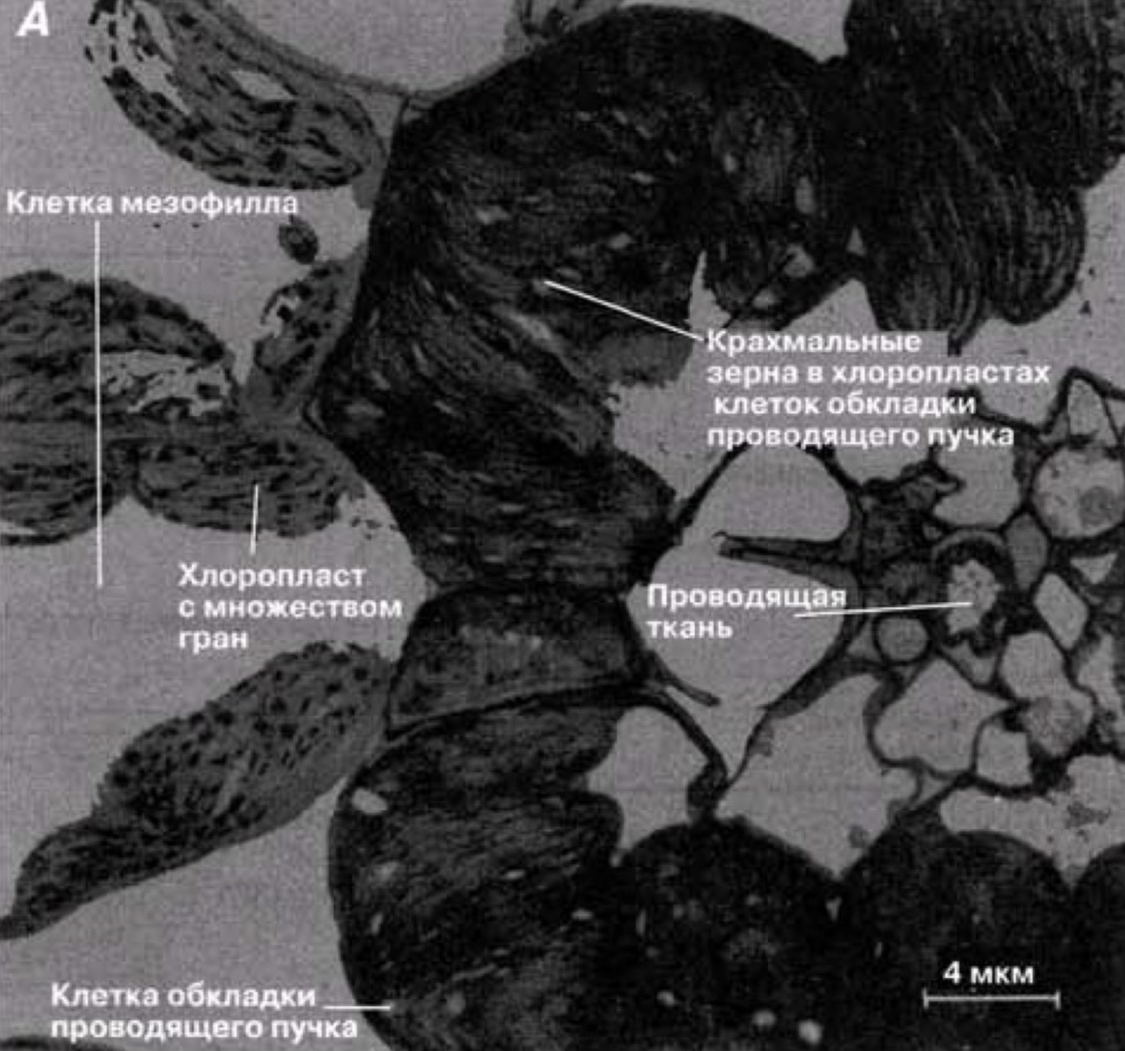
Начнем с того, что у С4-растений листья имеют характерную особенность строения: вокруг каждого проводящего пучка у них расположены два ряда клеток.
У клеток внутреннего кольца - обкладки проводящего пучка - хлоропласты по своей форме и строению отличаются от хлоропластов клеток мезофилла внешнего кольца. Хлоропласты в клетках мезофилла представляют собой высокоспециализированные органеллы, в которых протекают световые реакции фотосинтеза, а в хлоропластах клеток обкладки проводящего пучка протекают темновые реакции. Биохимические реакции проходящие на мембранах и строме хлоропластов как раз таки и представляют собой фотосинтез.
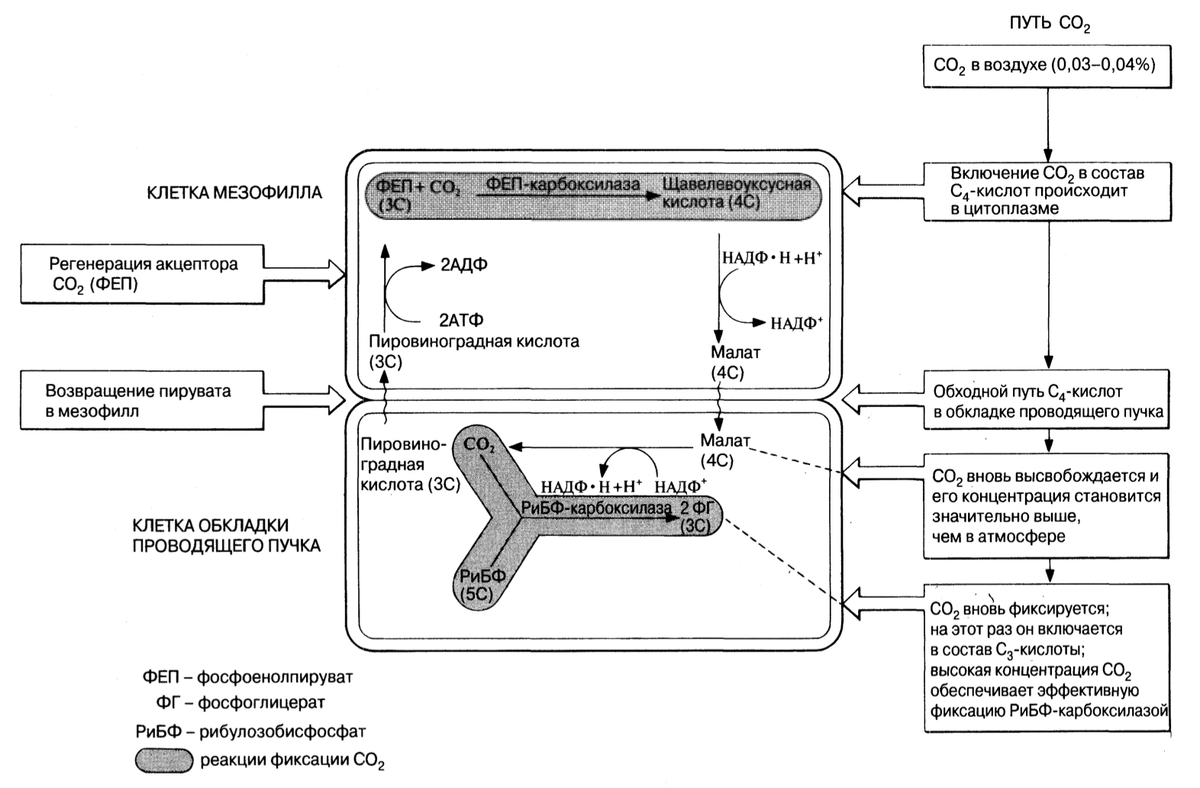
Путь Хэтча-Слэка
Рассмотрим теперь путь Хэтча-Слэка более подробно. По своей сути он предназначен для транспортировки диоксида углерода и водорода из клеток мезофилла в клетки обкладки проводящих пучков.А Из клеток обкладки проводящих пучков диоксид углерода высвобождается и поступает в обычный Сз-путь фотосинтетических превращений. Рассмотрим этот путь поэтапно.
1.Захват (фиксация) диоксида углерода в клетках мезофилла
Диоксид углерода фиксируется в цитоплазме клеток мезофилла. Механизм этого процесса приведен в следующем уравнении:
Акцептором диоксида углерода служит фосфоенолпируват (ФЕП) вместо РиБФ у С3-растений, а вместо фермента РиБФ-карбоксилазы у С4-растений участвует фермент ФЕП-карбоксилаза. Нужно отметить, что фермент ФЕП-карбоксилаза работает значительно более эффективно, чем фермент Сз-растений по двум причинам. Во-первых, ФЕП-карбоксилаза обладает большим сродством к диоксиду углерода, а во-вторых, ее работа не подвергается конкурентному ингибированию кислородом. Образовавшаяся щавелевоуксусная кислота далее превращается в малат, 4С-кислоту.
2.Малатный обходной путь (шунт)
Пройдя через плазмодесмы в клеточных стенках, малат попадает в хлоропласты клеток обкладки проводящего пучка, где он, соединяясь с диоксидом углерода, превращается в пируват (3С-кислоту). При этом выделяется водород, который используется для восстановления НАДФ. Обратите внимание, что в клетки мезофилла диоксид углерода и водород поступают извне, а затем в клетках обкладки проводящего пучка они вновь удаляются. Суммарным эффектом этих процессов является перемещение диоксида углерода и водорода из клеток мезофилла в клетки обкладки проводящего пучка.
3. Регенерация акцептора диоксида углерода
Пируват возвращается в клетки мезофилла, где в результате присоединения фосфатной группы от АТФ используется для регенерации ФЕП. На это расходуется энергия двух высокоэнергетических фосфатных связей.
Итоговый результат С4-пути
Итогом С4-пути является транспорт диоксида углерода и водорода из клеток мезофилла в хлоропласты клеток обкладки проводящего пучка с использованием двух высокоэнергетических фосфатных связей АТФ. Поскольку для обеспечения транспорта требуется энергия АТФ, работу данного механизма вполне можно сравнить с механизмом работы насоса.
Повторная фиксация диоксида углерода в клетках обкладки проводящего пучка и переход к С3-фотосинтезу.
В хлоропластах клеток обкладки проводящего пучка помимо пирувата образуется диоксид углерода и восстановленный НАДФ (говорилось выше при описание малатного шунта). Диоксид углерода затем вновь фиксируется РиБФ-карбоксилазой, участвующей в реакциях Сз-пути, а восстановленный НАДФ используется для восстановления ФГ(фосфоглицерата) (до сахара).
Особенности:
Основной особенностью С4-фотосинтеза является его энергозатратность: из-за повторной фиксации CO2. требует примерно в два раза больше энергии, чем C3-фотосинтез. Тогда данный процесс может показаться нелогичным, но это не так. Во-первых энергозатратность для процесса фотосинтеза не является лимитирующим фактором, тем более когда мы говорим о растениях тропических и субтропических поясов. Во-вторых лимитирующими фактороми для фотосинтеза можно считать как-раз таки фиксацию углекислого газа и, если говорить о засушливых регионах, наличие воды. И путь Хэтча-Слека прекрасно помогает обеспечить растение углекислым газом и также помогает снизить потери воды.
Высокая эффективность работы ФЕП-карбоксилазы обеспечивает высокую концентрацию CO2, что делает практически невозможным связывания РиБФ с кислородом (что в случаи классического фотосинтеза происходит относительно часто) и способствует улучшению работы фермента РиБФ-карбоксилазы, так как он начинает функционировать в среде с избытком субстрата. Также избыток диоксида углерода позволяет растениям достигать светового насыщения при более высоких значениях освещенности, то есть позволяет использовать более высокую освещеённость и температуру более эффективно.
Что касаемо сохранения воды, опять таки здесь помогает быстрая фиксация диоксида углерода. Обычные растения для того, чтобы терять меньше воды, должны уменьшать отверстия устьиц, что приводит к уменьшению площади проникновения углекислого газа. Но у С4-растений углекислый газ фиксируется настолько быстро, что по сравнению с С3-растениями они на каждую связанную молекулу CO2 теряют в 2 раза меньше воды.
Лично для меня это все напомнило один из законов диалектики «переход количества в качество»: большие затраты энергии, большая концентрация углекислого газа, приводят к росту эффективности использования энергии солнца, работы фермента и всего фотосинтеза в целом.
Заключение
C4-фотосинтез - это не альтернатива, а дополнение к «C3-пути», эволюционная адаптация, позволяющая растениям эффективно использовать солнечный свет, захватывать CO2 и сохранять воду в условиях высокой температуры и засухи. Он обеспечивает более высокую урожайность и устойчивость к неблагоприятным климатическим условиям, но требует дополнительных энергетических затрат.