Введение
Проторозаврии (лат.Protorosauria) – вымершая, возможно парафилетическая группа базальных архозавроморф, вероятно давшая начала всем остальным архозавроморфам, архозаврообразным и настоящим архозаврам. Впервые группа появляется ещё в поздней перми с таким родом животных как проторозавр, который был небольшим насекомоедом скрывавшимся в тени более крупных терапсид. Обитали проторозаврии на территории Европы, Азии и Сев. Америки вплоть до конца триаса (ретийский ярус).
В данной статье будут рассмотрены примитивные представители этой группы, которые уже постепенно начали стремиться к водной жизни, однако пока ещё были прикованы к суше.
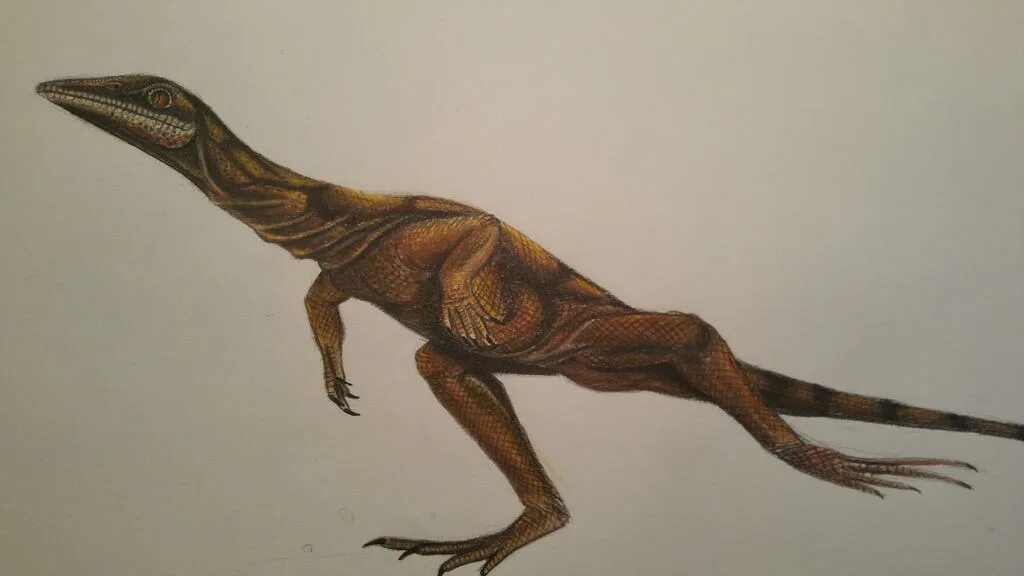
Чатковиелла
Чатковиелла (Czatkowiella) – вымерший род архозавроморфов проторозаврий, живший на территории Польши (Чатковица1) в оленёкском ярусе (поздний ранний триас). Типовой вид C.harae. Чатковиелла имела длинную шею, что является довольно распространенной чертой среди проторозаврий.
Джезайрозавр
Краткий очерк
Джейзайрозавр/джезарозавр/иесайрозавр (лат. Jesairosaurus) – вымерший род проторозаврий, живший на территории Алжира (провинция Иллизи) в раннем и среднем триасе (поздний оленёкский - анизийский). Типовой и единственный вид J.lehmani.
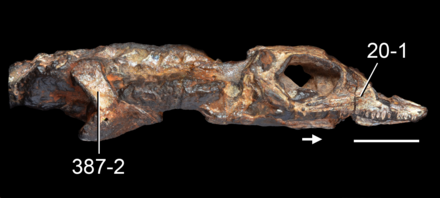
Описание
Череп
Кости рыла и небо
Череп низкий, с большими глазницами (глазными впадинами) и узкой и относительно короткой предглазничной областью (частью перед глазами). Точная форма ноздрей (носовых отверстий) неизвестна, но, судя по строению костей морды, они, вероятно, были полностью окружены большими, но узкими предчелюстными костями (парными костями, несущими зубы на кончике морды).Вентральный отросток (нижняя часть) каждой предчелюстной кости был длинным и содержал большое количество зубных ячеек. Хотя в правой предчелюстной кости голотипа сохранилось всего 6 зубов, там было место ещё для 9 или 10 зубов.
Каждая верхняя челюсть (зубная кость, расположенная сбоку от морды) имеет короткий передний и дорсальный отростки (выступающие вперёд и вверх части в передней части каждой верхней челюсти), а также длинный и массивный задний (задний) отросток, который образует большую часть нижнего края глазницы. Передняя часть верхней челюсти вогнутая, с отверстием над третьим зубом. В каждой верхней челюсти было от 20 до 21 зуба. Передний край каждой глазницы был образован лобной костью, а между каждой лобной костью и верхней челюстью располагалась тонкая слёзная кость. Слёзная кость, вероятно, не образовывала часть границы глазницы или ноздрей.
Нёбные кости в верхней части ротовой полости в передней части морды плотно прилегали к верхнечелюстным костям. Некоторые кости в передней части нёба (вероятно, сошники) также были покрыты мелкими зубцами. Зубцы также присутствовали в задней части нёба, вероятно, на крыловидных костях и задних частях нёбных костей. Межптеригоидные полости (большие отверстия по бокам нёба) были длинными и тонкими, а подглазничные отверстия (отверстия в задней части нёба, через которые проходят мышцы) — большими.
Заглазничная область
Задние нижние углы глазниц образованы скуловыми костями, которые простираются до нижней части глазничной области черепа (за глазами). Каждая скуловая кость имела длинный и похожий на шпору задний отросток, который доходил до чешуйчатой кости в задней части черепа. Однако этот задний отросток простирался над каждым нижним височным отверстием — отверстием в боковой части задней части черепа. Таким образом, отверстие остаётся полностью открытым снизу, в то время как у многих других рептилий подъязычная кость закрывает отверстие снизу.
Прямо над каждой скуловой костью находится треугольная скуловая кость. Тонкий дорсальный отросток каждой скуловой кости образует часть переднего края каждой верхней височной фенестры — большого круглого отверстия в верхней части задней части черепа. Передний край каждой скуловой кости соприкасается с довольно крупной заднелобной костью, которая образует часть как заднего края глазницы, так и переднего края верхней височной фенестры. За задним концом каждой скуловой кости находится ромбовидная чешуйчатая кость, которая образует нижний край верхней височной фенестры. Вогнутый задний край каждой чешуйчатой кости сочленяется с выпуклым передним краем каждой квадратной кости, образуя прочное и негибкое соединение «шип-паз». Каждая квадратная кость прочная и направлена вниз и назад, но нет никаких доказательств того, что у иесайрозавра были квадратно-скуловые (кости, соединяющие скуловую и квадратную кости) кости.Кроме того, у квадратной кости отсутствует заметно вогнутый задний край и выступающий вперёд передний край, в отличие от лепидозавроморфов.
Крыша черепа и черепная коробка
Лобные кости, занимающие часть свода черепа между глазами, имеют прямоугольную форму и образуют большую часть верхнего края глазниц.Теменные кости, расположенные за лобными и между верхними височными отверстиями, меньше лобных и имеют боковые (направленные наружу) выступы, которые выступают вниз и образуют задний край верхних височных отверстий.Передняя часть теменных костей сужается внутрь над глазницами, и у некоторых экземпляров (таких как ZAR 07) можно увидеть небольшое шишковидное отверстие, полностью закрытое этой частью теменных костей. Шишковидное отверстие — это отверстие в середине черепа, в котором у некоторых современных рептилий находится сенсорный орган, который иногда называют «третьим глазом».
Затылочная кость, часть черепа, расположенная непосредственно над большим затылочным отверстием (отверстием в черепе для спинного мозга), большая и заметная сверху, но не соприкасается с теменными костями. Затылочные отростки сосцевидных костей (внутренних слуховых косточек со стороны большого затылочного отверстия) достаточно широкие, чтобы достигать чешуйчатых, квадратной и латеральных частей теменных костей по бокам головы. Базиокципитальная кость непосредственно под большим затылочным отверстием имеет небольшой киль вдоль нижнего края. Затылочный мыщелок, продолжение базиокципитальной кости, соединяющееся с позвонками, расположен на черепе дальше, чем сустав между черепом и нижней челюстью.
Нижняя челюсть и зубы
Нижние челюсти (мандибулы) прямые и тонкие, образованы несущими зубы зубными костями спереди и сфеноидальной и угловатой костями сзади. Передний конец нижней челюсти слегка изогнут вниз и внутрь. Зубы иесайрозавра заострённые и слегка изогнутые, хотя они также конические (особенно в верхней челюсти) и лишь слегка уплощённые с боков. Кроме того, зубы были субтекодонтными (также известными как плевротекодонтными). Это означает, что зубы располагались в лунках внутри длинной бороздки, ограниченной костными стенками, при этом губная (внешняя) стенка была выше язычной (внутренней).
Позвоночник и ребра
У иесайрозавра было 9 шейных (позвоночных) позвонков, и у него была необычно короткая шея по сравнению со многими другими базальными архозавроморфами, в частности с причудливыми танистрофеями, с которыми он был связан.Остистые отростки, выступавшие из верхней части каждого позвонка, были низкими и узкими, за исключением высокого остистого отростка оси (второго шейного позвонка). У некоторых экземпляров все остистые отростки были наклонены вперёд, в то время как у других экземпляров такое положение имели только последние несколько позвонков, а остальные поднимались вертикально. Это пример индивидуальной вариации в пределах рода. Несмотря на плохую сохранность, шейные рёбра у экземпляров иесайрозавра были длинными и тонкими для всех шейных позвонков, кроме осевого, у которого они были короткими.
У него было по меньшей мере 15 спинных (дорсальных) позвонков. Остистые отростки этих позвонков также были низкими и узкими, хотя их высота увеличивалась по направлению к задней части тела. У некоторых особей средние дорсальные позвонки имеют дополнительные отростки, направленные вперёд или назад, у основания остистых отростков. Центры (основные тела) первых нескольких спинных позвонков выше, чем шире, и имеют округлую форму, при этом каждый из них имеет вогнутый нижний край. У некоторых также есть неглубокие впадины по бокам. Позвонки становятся шире и прочнее к задней части тела. У взрослых особей этого рода в позвонках отсутствует канал для спинного мозга. Также присутствуют крупные пластинчатые зигапофизы (суставы между позвонками), наряду с диапофизами и направленными вперёд парапофизами (различные типы рёберных суставов сбоку и спереди каждого позвонка соответственно). С другой стороны, у этого рода полностью отсутствуют межцентральные кости — маленькие кости, расположенные между центрами позвонков у некоторых четвероногих. Большинство спинных рёбер длинные и выступают наружу и назад, за исключением последних трёх, которые короткие и слегка направлены вперёд. Три ряда длинных Гастралии (брюшные рёбра) также присутствуют.
Присутствовали 2 крестцовых (тазовых) позвонка с высокими и тонкими остистыми отростками. Первые несколько шейных (хвостовых) позвонков также имели высокие и тонкие остистые отростки, хотя большая часть хвоста неизвестна. Хвостовые рёбра короткие, направлены назад и срастаются с позвонками.
Грудной отдел и передние конечности
Каждая лопатка (плечевая кость) и клювовидно-плечевая кость (плечевой пояс) срастаются в лопаточно-клювовидно-плечевую кость, хотя их всё ещё можно различить по уплощённой области на кости. Тонкая лопатка расширяется в веерообразную структуру по мере продвижения вверх и назад. Каждая клювовидно-плечевая кость сохраняется в виде большой и широкой пластины, которая расширяется к задней части и имеет отверстие (клювовидно-плечевое отверстие). При взгляде спереди две стороны грудного пояса образовывали угол в 80 градусов. Вдоль переднего края каждой ключичной кости располагалась стержнеобразная ключичная кость, а между двумя ключичными костями находилась узкая Т-образная межключичная кость. За грудным поясом располагались возможные овальные грудинные пластины. Эти пластины представляют собой костные компоненты грудины (передней части груди), которая у большинства рептилий полностью хрящевая, а у птиц — полностью костная.
Каждая плечевая кость (верхняя часть руки, единственная полностью сохранившаяся часть передних конечностей) узкая в средней части, но расширена на проксимальном (ближнем к телу) и дистальном (дальнем от тела) концах. Проксимальный конец является самой широкой частью, а на дистальном конце имеется большая выемка вдоль переднего края, образующая крючкообразную структуру. Как надмыщелковое, так и подмыщелковое отверстия (два отверстия на дистальном конце плечевой кости) полностью закрыты. Также сохранились проксимальные концы лучевой и локтевой костей (нижняя часть руки), что указывает на то, что это были тонкие кости.
Таз и задние конечности
Каждая подвздошная кость (верхняя пластина таза) была очень похожа на таз пролацерты и трилофозавра. Каждая подвздошная кость имела треугольную форму и была разделена на две части толстым гребнем, который образовывал верхний край большой вертлужной впадины (тазовой ямки). Нижняя часть каждой стороны таза состояла из «лобково-седалищной пластины», образованной слиянием выступающей вперёд лобковой кости и выступающей назад седалищной кости. Седалищная кость не выступает дальше задней части подвздошной кости. Две лобково-седалищные кости по обеим сторонам таза срастаются внизу под углом 80 градусов при виде спереди.
Бедренная кость (тазовая кость) была прямой и похожей на стержень, с вогнутой и костлявой головкой, хотя многие детали неизвестны из-за того, что кость была неполной во всех образцах. Большеберцовая и малоберцовая кости (кости нижней части ноги) также были длинными, прямыми и тесно связанными. В целом задние конечности, по-видимому, были длиннее передних, что, возможно, позволяло животному какое-то время бегать на двух ногах. Пястные кости (кости стопы) сохранились плохо, но пяточной кости (камню пятки), по-видимому, не хватало пяточного бугра (большой костной выпуклости сбоку кости).
Ромбофолис
Ромбофолис (Rhombopholis) –род проторозавров из анизийского яруса среднего триаса (~245 млн. лет назад) Англии. Типовой вид R.scutulata был обнаружен в карьере Лимингтон, недалеко от Уорика, что относится к формации Броумсгровского песчаника.
Ромбофолис известен по нескольким различным образцам. WARMS Gz10, блок песчаника, содержащий позвонки и кости конечностей двух разных особей, а также рыбью чешую и позвонки, был назначен лектотипом рода. К другим образцам относятся WARMS Gz21 ( проксимальная часть левой бедренной кости), WARMS Gz4714 (левая подвздошная кость) и CAMSM G.343 (позвонок). Ромбофолис не имеет отличительных особенностей, но считается «пролацертиформным» (устаревший термин для обозначения базального архозавроморфа) из-за низких нервных отростков.
Более мелкая особь, присутствующая в WARMS Gz10, представлена тремя шейными позвонками и частью бедренной кости. Центры позвонков широкие, но сужены посередине, в то время как нервные отростки длинные и низкие. Бедренная кость длинная и прямая, с выпуклым передним краем на проксимальном конце.
Более крупная особь в WARMS Gz10 представлена неполным хвостовым позвонком, ребром, проксимальным концом правой бедренной кости и несколькими метаподиями. Ребро прямое, но на своем проксимальном конце расширяется в два сочленения. Бедренная кость имеет продольный гребень и похожа на таковую у танистрофея и малеризавра. Предполагаемая плюсна имеет гребень на проксимальном конце, плоскую ножку и широкий дистальный конец с небольшим углублением. Предполагаемая пястная кость плоская и слегка изогнутая, с более широким проксимальным концом. Другие экземпляры ромбофолиса по размеру близки к этой особи.
CAMSM G.324 напоминает поясничные позвонки макрокнема. WARMS Gz21 имеет широкое заднее расширение и узкое переднее расширение. По сравнению с Macrocnemus, общая длина WARMS Gz21 составляет 134 миллиметра, в то время как длина бедренной кости более крупного экземпляра WARMS Gz10 составляет 144 миллиметра. Несмотря на то, что ромбофолис крупнее чем макрокнем, он всё равно меньше, чем несколько видов танистрофеид. WARMS Gz4714 имеет короткий передний отросток и широкую вертлужную впадину с сильно утолщёнными краями.
Мегакнем
Мегакнем (Megacnemus) – род проторозавров живший на территории Польши в среднем триасе (245-240 миллионов лет назад). Типовой вид M.grandis известен лишь по единственной бедренной кости длина которой составляет 20 сантиметров, что позволяет предположить о крупных размерах мегакнема сравнивого с архозавроморфом-вритраминозавром (V. dzerzhinskii) из раннего триаса России.
Предполагаемая бедренная кость мегакнема отличается от костей других базальных архозавроморфов тем, что она довольно прочная и прямая, в отличие от более сигмовидно-изогнутых бедренных костей проторозавра, макрокнема и танистрофеид. Однако у неё есть некоторые сходства с плечевой костью диноцефалозавра и макроцении, такие как расширенные дистальный и проксимальный концы, прямой преаксиальный край и вогнутый постаксиальный край, хотя концы кости менее округлые, чем у этих таксонов. Кость также напоминает плечевую кость бореоприцеи, хотя она более уплощённая.
Малютинизух
Малютинизух/малутинизух (Malutinisuchus) — вымерший род архозавроморфов проторозаврий из раннего и среднего (245 млн. лет назад) триаса России. Типовой вид M. gratus. Малутинизух также известен по отложениям среднего триаса ладинского яруса в Букобайской и Рассыпной впадинах в Оренбургской области, Россия. В России отложения этого возраста относятся к Букобайскому горизонту.
В подборку так же мог входить род эксиллизухов (Exillisuchus), но его положение в классификации остаётся спорным.