Механические свойства яичниковой ткани изменяются в течение репродуктивной жизни женщины и играют решающую роль в активации фолликулов. Понимание этих свойств открывает новые возможности для лечения бесплодия и разработки методов in vitro фолликулогенеза. Хотя рост от вторичных до антральных фолликулов хорошо изучен, инициация фолликулогенеза остается малоизученной.
Данная статья является адаптированным переводом оригинальной статьи "Biomechanical properties of ovarian tissue and their impact on the activation of follicular growth: a narrative review"

Введение в биомеханику яичника
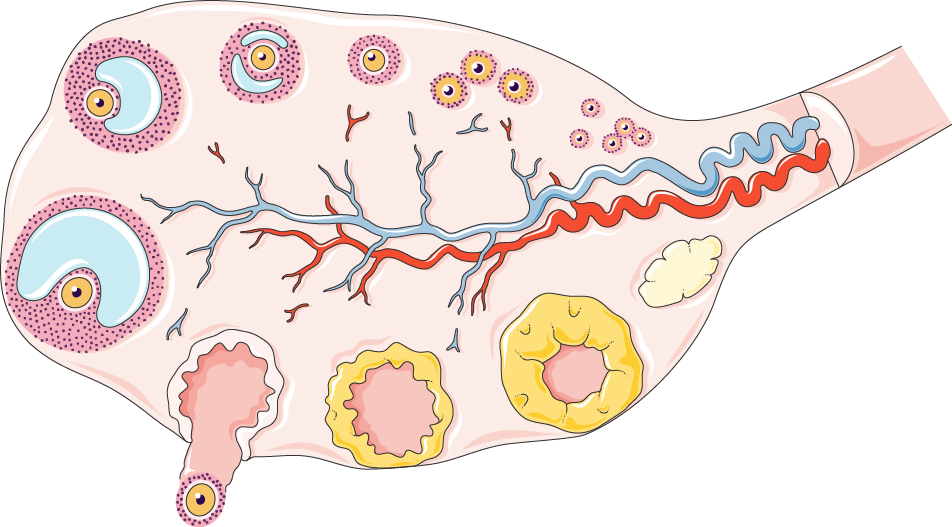
Основная функция яичника - обеспечить овуляторный фолликул и выпустить зрелый ооцит, способный к оплодотворению. Самые молодые фолликулы, называемые "примордиальными фолликулами", расположены в коре яичника. Примордиальный фолликул состоит из ооцита, окруженного одним слоем уплощенных гранулезных клеток.
После активации примордиальный фолликул растет и превращается в первичный (преантральный) фолликул. Затем он превращается во вторичный (антральный) фолликул, образуя многочисленные жидкостные компартменты внутри скопления гранулезных клеток.
С детства каждый месяц активируется множество фолликулов, что приводит к постепенному уменьшению пула фолликулов на протяжении жизни женщины и к менопаузе. Рекрутинг фолликулов регулируется очень точно, чтобы поддерживать долгосрочный баланс между пулами покоящихся и растущих фолликулов, обеспечивая производство одного зрелого ооцита в месяц и предотвращая преждевременное истощение овариального резерва. В активации части пула покоящихся фолликулов каждый цикл участвуют несколько сигнальных путей, таких как пути PI3K-Akt, Hippo и mTOR.
Ряд исследований показал, что рост фолликулов чувствителен к микроокружению. Это подчеркивает необходимость понимания физических и механических свойств ткани яичника, которые необходимы для контроля in vitro фолликулогенеза.
Внеклеточный матрикс яичника: состав и функции
Яичник - это высокодинамичная ткань, которая претерпевает циклы роста и овуляции на протяжении всей репродуктивной жизни. Эти изменения возможны благодаря ремоделированию эластиновых и коллагеновых волокон внеклеточного матрикса (Далее - ВКМ), которые составляют субстрат, поддерживающий клеточные процессы, возникающие в результате комбинации механических и химических сигналов.
ВКМ регулирует выживание, адгезию, пролиферацию и дифференцировку гранулезных клеток, помогает поддерживать сферическую форму фолликулов. Более того, он также регулирует продукцию прогестерона начиная со стадии вторичного фолликула и концентрацию эстрадиола. Соответственно он регулирует созревание ооцита.
ВКМ окружает фолликулы с примордиальной стадии и находится по всей коре и мозговому веществу, обеспечивая жесткую или эластичную механическую поддержку яичнику. Он представляет собой биомакромолекулярную сеть, в которой находятся фолликулы и стромальные клетки, и состоит из воды. Исследования протеома коры яичника выявили более 120 белков, которые структурируют, изменяют форму и регулируют ВКМ, часть наиболее важных из них представлена в Таблице 1.
Изменения в структуре ВКМ яичника влияют на механические свойства. Эти механические свойства изучались и количественно определялись в яичниках различных видов с использованием различных методов.
В литературе механические свойства тканей (и даже клеток) часто описываются с точки зрения эластичности (значение модуля упругости), вязкоупругости и жесткости образца. Эти физические свойства могут варьироваться в яичнике, но также и внутри фолликулов. Материал считается идеально упругим, если он мгновенно деформируется под напряжением и возвращается к своей исходной форме, когда напряжение удаляется.
Эластичность изотропных материалов полностью количественно определяется двумя независимыми упругими дескрипторами: модулем Юнга (E) и модулем сдвига (G).
Вязкость измеряет сопротивление жидкости (жидкости) деформации либо сдвиговым напряжением, либо растягивающим напряжением. Это результат диффузии и взаимодействия молекул внутри аморфного материала, способного течь. Некоторые материалы проявляют как вязкие, так и упругие характеристики (вязкоупругость).
Несколько авторов обнаружили, что кора яичника более упругая, а мозговое вещество более вязкое. При низких деформациях в динамическом механическом анализе, где напряжение и деформация являются синусоидами, яичник проявляет вязкоупругие свойства. (Рисунок 1).
Механические свойства яичника
Яичник состоит из двух ключевых зон:
- Кора — плотная и жесткая, сохраняет "спящие" примордиальные фолликулы. В коре много коллагена и гиалуроновой кислоты, что создает жесткий каркас.
- Мозговое вещество — более мягкое, поддерживает рост фолликулов до зрелости. Мозговое вещество содержит меньше этих компонентов, обеспечивая гибкость для роста фолликулов.
При изучении жесткости ткани используют 2 основных метода: Атомно-силовая микроскопия (AFM) показывает, что края яичника жестче (7 кПа), чем центр (2-3 кПа), а ультразвук и МРТ - хоть и неинвазивные методы, но дают противоречивые данные. Например, у животных кора мягче мозгового вещества, а у людей различий может не быть.
Что происходит во время роста фолликулов? Изменения в матриксе:
Толстые коллагеновые волокна заменяются тонкими, что облегчает рост.
Эластин уменьшается, а гликозаминогликаны остаются стабильными. Роль жесткости: Жесткая среда (например, 2% гидрогель) сохраняет фолликулы в покое. Мягкие условия (0.25% гидрогель) стимулируют рост и созревание ооцитов.
Практическое применение: Искусственные матрицы - гидрогели с регулируемой жесткостью имитируют естественную среду яичника, улучшая культивирование фолликулов in vitro. Лечение бесплодия - понимание механики ткани помогает разрабатывать методы сохранения фертильности, например, при химиотерапии. Баланс между жесткостью и эластичностью яичника критичен для сохранения фолликулярного резерва и своевременного созревания яйцеклеток. Нарушения этого баланса (например, при старении или поликистозе) ведут к бесплодию, а их коррекция открывает новые пути лечения.
Было показано, что ремоделирование ВКМ, окружающего фолликулы, необходимо для их роста (Рисунок 2).
Механическая регуляция роста фолликулов: как клетки «чувствуют» жесткость среды
Клетки фолликулов воспринимают не только химические вещества, но и физические свойства окружающей ткани. Этот процесс, называемый механотрансдукцией, превращает механические воздействия (например, жесткость) в биохимические сигналы.
- «Антенны» клетки: Интегрины (белки-«крючки»), реснички и ионные каналы улавливают изменения в структуре внеклеточного матрикса (ВКМ).
- Сигнальные пути: Активируются внутриклеточные каскады (например, PI3K-Akt и Hippo), которые влияют на рост или покой фолликулов.
1. Путь PI3K-Akt. Активация роста: При снижении жесткости ВКМ белок FOXO3 перемещается из ядра в цитоплазму, снимая блок с роста фолликулов. Пример: Деградация коллагена ферментами «освобождает» фолликулы, запуская их развитие.
2. Путь Hippo. Сохранение покоя: В жесткой коре яичника активируется Hippo, блокируя рост фолликулов. При снижении жесткости (например, во время полового созревания) Hippo «выключается», позволяя фолликулам расти.
3. Роль белков YAP/TAZ. Механические сенсоры: Эти белки перемещаются в ядро при снижении жесткости, запуская рост клеток. Пример: При растяжении ткани YAP/TAZ активируют гены, необходимые для развития фолликулов.
Стимуляция роста фолликулов in vitro
Два подхода к активации фолликулов
- Биохимический метод:
Используются активаторы пути PI3K-Akt (стимулируют рост) и ингибиторы PTEN (блокируют «тормоза» развития). Пример: Трансплантация обработанной ткани пациентам с преждевременной недостаточностью яичников привела к успешным беременностям. - Механический метод:
Растяжение ткани на 10% от исходного размера перед культивированием.
Результат: Увеличение доли растущих фолликулов (например, с 2.9% до 10.8% через 8 дней).
Подготовка ткани происходит следующим образом: удаление избыточной стромы и сохранение коркового слоя с примордиальными фолликулами. Форма образцов важна: полоски ткани растут лучше, чем кубики, благодаря меньшей плотности стромы. При растяжении происходит ремоделивание внеклеточного матрикса - снижение коллагена (жесткость ↓) и рост эластина (эластичность ↑).
Улучшение доступа кислорода и факторов роста к фолликулам. Происходит активация сигнальных путей: нарушение баланса между путями PI3K-Akt и Hippo запускает рост фолликулов. Фрагментация ткани может имитировать естественные условия для миграции фолликулов в мозговое вещество.
Механические свойства внеклеточного матрикса на протяжении репродуктивной жизни
Ткань яичника с возрастом претерпевает множество изменений: до полового созревания (препубертат) ВКМ очень жесткий из-за плотной сети тонких коллагеновых волокон. Ткань мало эластина, что ограничивает рост фолликулов. Во время репродуктивного возраста коллаген перестраивается в толстые пучки, образуя более крупные поры, это облегчает доступ питательных веществ и гормонов к фолликулам. Содержание эластина и глюкозаминогликана увеличивается, делая ткань мягче и эластичнее. Оптимальный баланс жесткости и гибкости поддерживает созревание яйцеклеток. Во время менопаузы жесткость резко возрастает из-за накопления коллагена I и III типов (фиброз). Снижение эластина и гиалуроновой кислоты нарушает структуру ВКМ. И как результат происходит истощение фолликулярного резерва и прекращение овуляции (Рисунок 4).
Выводы
Механические свойства внеклеточного матрикса (ВКМ) яичника играют важную роль в регуляции фолликулогенеза и фертильности. Исследования показывают, что жесткость тканей напрямую влияет на способность фолликулов активироваться и развиваться. Например, мягкая среда с низким содержанием коллагена и высоким уровнем эластина создает оптимальные условия для роста примордиальных фолликулов, тогда как жесткий матрикс, характерный для стареющего или патологически измененного яичника, подавляет этот процесс. При старении накопление коллагена I и III типов приводит к фиброзу, что снижает овариальный резерв и нарушает овуляцию. В менопаузе деградация эластина и гиалуроновой кислоты усугубляет эти изменения, делая ткань менее функциональной.
Механические методы, такие как растяжение ткани на 10%, открывают новые возможности в репродуктивной медицине. Эксперименты, проведенные рядом исследователей, продемонстрировали, что контролируемая деформация коры яичника активирует сигнальные пути PI3K-Akt и Hippo, стимулируя переход фолликулов из «спящего» состояния в активное. Этот подход уже позволил получить зрелые ооциты in vitro, что особенно важно для пациенток, сохраняющих фертильность перед химиотерапией. Однако методы механической активации требуют осторожности: чрезмерное растяжение может вызвать необратимую деформацию, а длительное культивирование ткани повышает риск экспрессии опухолевых генов.
Клиническое применение этих наработок разнообразно. Например, при преждевременной недостаточности яичников (ПНЯ) трансплантация аутологичной коры после механической обработки восстановила фолликулогенез у половины пациенток в одном из исследований, приведя к двум успешным беременностям. При синдроме поликистозных яичников (СПКЯ), где плотный коллагеновый матрикс блокирует рост фолликулов, комбинация механического растяжения и биохимических ингибиторов может преодолеть ановуляцию.
Перспективы направления связаны с разработкой персонализированных терапий. Создание гидрогелей, имитирующих «молодой» ВКМ, улучшает культивирование фолликулов in vitro. Ингибиторы коллаген-синтетазы могут замедлить фиброз, продлевая репродуктивный период. Интеграция искусственного интеллекта для анализа механических свойств ткани способна оптимизировать прогнозирование ответа на лечение.