Перевод из Withrow & MacEwen’s Small Animal Clinical Oncology 6th
Представление о том, что мутации, которые накапливаются стохастически во время нормальной репликации клеток, вызывают злокачественную трансформацию, поднимает важный вопрос: почему более крупные животные, имеющие больше клеток, не более уязвимы для рака? Этот вопрос, первоначально заданный сэром Ричардом Пето, обычно называют парадоксом Пето (на уровне вида заболеваемость раком, по-видимому, не коррелирует с количеством клеток в организме). Ответ заключается в эволюции и естественном отборе. У млекопитающих с экстремальными размерами было выявлено несколько механизмов защиты от рака. Слоны являются крупнейшими из ныне живущих наземных млекопитающих, в генетике слонов защита от рака, по-видимому, связана с повышенной активностью гена-супрессора опухолей TP53, который претерпел множественные дупликации после отделения от общего предка, который дал начало даманам и ламантинам, но предшествовал диверсификации в мастодонтов, мамонтов и современных слонов. Киты являются крупнейшими ныне живущими животными, и в генетике гренландских китов, например, варианты или изменения нескольких генов, по-видимому, обеспечивают защиту от рака и старения. Однако у обыкновенного малого полосатика развилась альтернативная адаптация, которая привела к меньшему количеству микросателлитов в геномных областях вблизи протоонкогенов и генов-супрессоров опухолей, где кумулятивные мутации могут привести к повышенному риску рака. Как иллюстрируют эти примеры, адаптивные решения, которые обеспечивают большой размер и долголетие, вероятно, являются уникальными и специфичными для эволюционной истории каждого вида (рис. 1.2A).
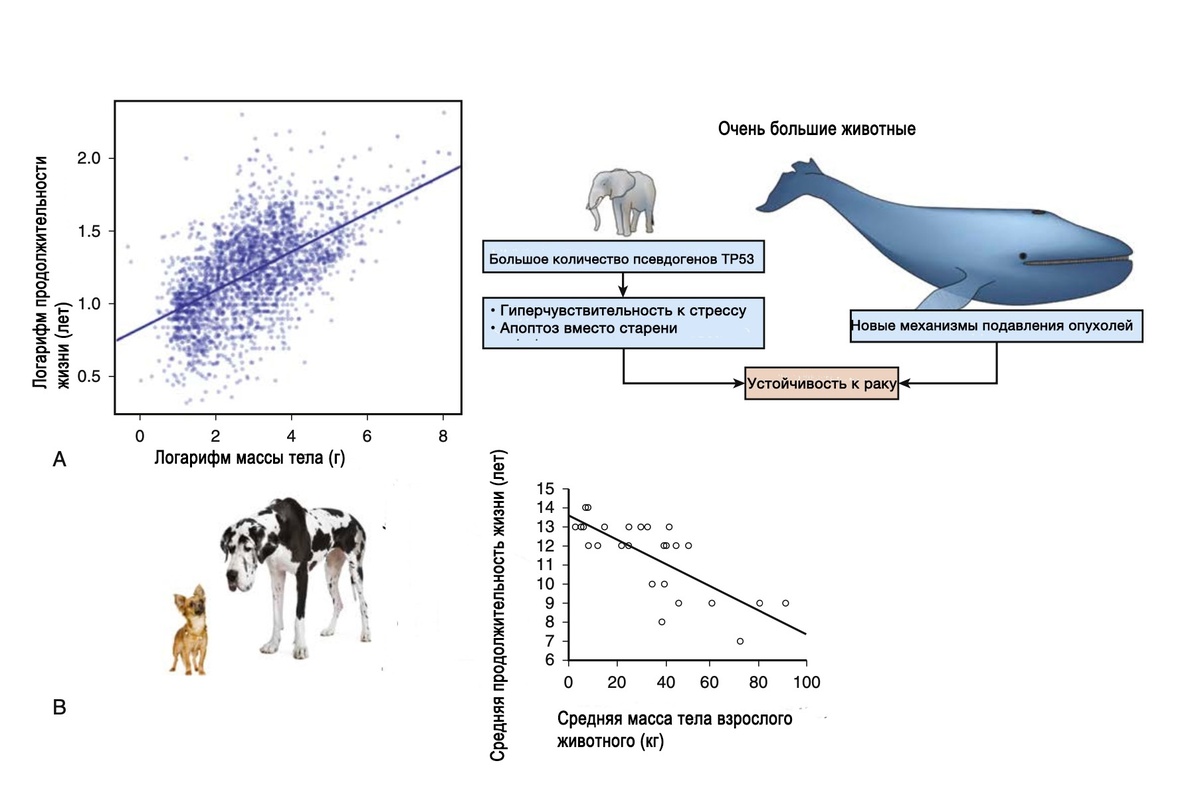
• Рис. 1.2 Размер тела и продолжительность жизни. (A) Связь между массой тела (г) и продолжительностью жизни (годы) среди 2556 позвоночных. На левой панели синяя линия показывает линейную регрессию между логарифмом (масса тела) и логарифмом (продолжительность жизни), R2 = 0,32. Правая панель иллюстрирует потенциальные противораковые механизмы у крупнейших млекопитающих: слонов и китов. У слонов развилось несколько копий гена TP53 (псевдогены), которые связаны с повышенным апоптотическим ответом. Противораковые механизмы у крупнейших млекопитающих, китов, пока не известны, но они не включают дупликации TP53. (B) Связь между массой тела и продолжительностью жизни у 32 различных пород собак. Разнообразие размеров и продолжительности жизни среди пород собак примечательно, но также хорошо известно, что более крупные породы, как правило, живут недолго по сравнению с мелкими породами.
Нормой эволюции млекопитающих является то, что большой размер коррелирует с продолжительностью жизни. Это согласуется с селективным давлением, которое в противном случае привело бы к снижению энергетических затрат, необходимых для достижения больших размеров. Однако у домашних собак эта тенденция противоположна, у них большой размер тела связан с более короткой продолжительностью жизни и, возможно, с более высоким уровнем некоторых заболеваний, включая рак (см. рис. 1.2В). Точные причины этого остаются неясными, но собаки представляют собой уникальную естественную модель для изучения взаимосвязей между возрастом, массой тела и риском заболеваний в условиях, когда искусственный отбор вытеснил естественный. В частности, естественное селекционное давление у собак было заменено искусственным отбором с момента первоначального одомашнивания примерно 10 000-25 000 лет назад. Изменение спроса на форму, а не на функциональность привело к созданию более 400 пород за последние 300-400 лет. Этот искусственный отбор, обычно по одному или нескольким фенотипическим признакам, дает мало шансов на адаптацию по всему остальному геному. Следовательно, риск мутаций, связанных с нормальными процессами репликации клеток во время развития, роста и поддержания их во взрослом состоянии, повышается у крупных собак (больше клеток), что позволяет объяснить непропорционально высокий риск некоторых видов рака, таких как аппендикулярный остеопороз, манипуляциями с их геномной пластичностью с помощью экстремального отбора из-за размера. Это также согласуется с тем фактом, что общий риск развития аксиальной остеосаркомы у собак аналогичен тому, который наблюдается у других видов, что объясняется влиянием размера и функциональных/механических нагрузок на кости. Повышенный риск развития аппендикулярной остерсаркомы у крупных и гигантских собак может быть объяснен, по крайней мере частично, тем фактом, что для создания и поддержания крупных костей требуется больше клеточных делений, особенно в связи с тем, что костные ткани подвергаются постоянному ремоделированию. Каждый раунд репликации остеобласта, в свою очередь, вносит свой вклад в его мутационную нагрузку и потенциальную трансформацию. Поздний возраст начала заболевания у собак согласуется с хроническим отбором клеток, которые накапливают критический набор мутаций. У людей остеосаркома относится к числу видов рака, при которых R-фактор может объяснить практически весь риск; кроме того, была выявлена связь, которая показывает, что остеосаркома чаще встречается у детей в более высоких процентилях роста для их возраста.
Учет риска аппендикулярной остеосаркомы (или других типов рака) стохастически как функции риска репликации оставляет без ответа еще один важный вопрос: почему риск для некоторых пород собак оказывается выше (или ниже), чем ожидалось, исходя из их общего размера? На этот вопрос есть частичные ответы, и, опять же, имеются достоверные данные по остеосаркоме. Что касается риска, связанного с породой, то, по-видимому, на риск развития остеосаркомы влияют многочисленные наследственные факторы. Данные общегеномного ассоциативного исследования (GWAS) трех пород собак, относящихся к группе высокого риска, указывают на то, что закономерности наследственного риска развития остеосаркомы являются сложными и не полностью понятными. Однако селекция, особенно для получения крупного размера, по-видимому, обогатила аллели риска, которые в настоящее время фиксированы в определенных популяциях. Фиксированные аллели, ассоциированные с риском, характерны не только для остеосаркомы; они также были связаны с риском развития рака молочной железы у собак, плоскоклеточного рака пальцев у собак., и других видов рака (обсуждаются далее в этой главе).
У людей GWAS выявил два локуса, связанных с наличием остеосаркомы, и один, отдельный локус в гене NFIB, связанный с наличием метастазов при постановке диагноза. Аллели риска для людей и собак не расположены в ортологичных областях генома. Таким образом, оказывается, что относительно быстрое достижение большого размера, а не специфичные для породы (или индивидуальные) признаки в зародышевой линии, является подавляющим фактором риска остеосаркомы. Таким образом, разумно заключить, что парадокс Пето возникает в результате барьеров естественного отбора, и что когда такие барьеры устраняются и естественная продолжительность жизни организма увеличивается, как в случае с собаками, парадокс исчезает, раскрывая подавляющее влияние ошибок репликации ДНК на индивидуальный риск рака.
Приведенные примеры подчеркивают, что риск рака и прогрессирование имеют как общие, так и уникальные черты у разных видов в животном мире. Это создает возможности для изучения естественной истории рака в спонтанной обстановке. Однако важно избегать существенных ловушек, которые возникают из-за предположений об эквивалентности, когда ее нет. В следующем разделе этой главы рассматриваются отличительные признаки рака, которые являются общими практически для каждого вида рака. Заключительный раздел главы посвящен недавним исследованиям, которые выдвигают собак-компаньонов в качестве моделей для понимания сложной генетики рака посредством использования современных технологий и осторожной, преднамеренной интерпретации данных.