Клеточный цикл
Клеточный цикл (жизненный цикл клетки) – промежуток времени от момента возникновения клетки в результате деления исходной материнской до ее собственного деления или гибели.
Обязательным компонентом клеточного цикла большинства соматических клеток является митотический цикл, который включает интерфазу и митоз. У ряда авторов понятия «митотический» и «клеточный» циклов совпадает.
Интерфаза – период подготовки клетки к делению.
Характеристика интерфазы
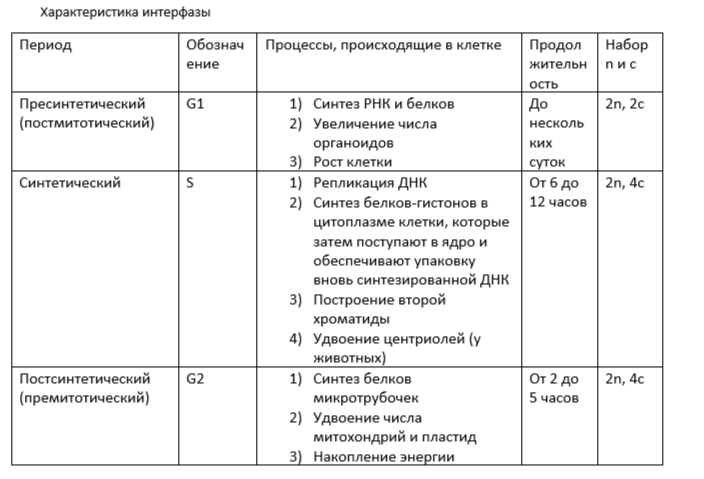
В период G1 синтезируются особые белки – активаторы S-периода. Они обеспечивают достижение клеткой определенного порога точки R (рескрипции, ограничения), после которой наступает следующий S-период. Если клетки не достигнут точки R, она выходит из цикла и вступает в период репродуктивного покоя G0, для того чтобы:
1) Дифференцироваться и выполнять свои функции,
2) Выжить в условиях недостатка питательных веществ,
3) Осуществить репарацию поврежденной ДНК.
Клетки одних тканей при соответствующей стимуляции способны вновь возвращаться из периода G0 в клеточный цикл (в митотический); а клетки других тканей эту способность утрачивают.
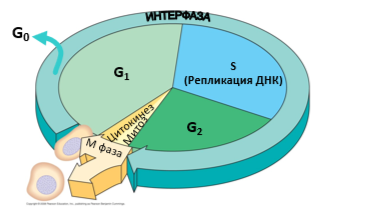
Митоз (непрямое деление)
греч. Mitos – нить
Этот процесс понимают в широком и более узком смысле слова. В широком смысле митоз – деление соматической клетки, сопровождающееся образованием веретена деления и равномерным распределением генетического материала в дочерние клетки. Митоз следует за периодом G2 и длится в среднем 1-3 часа. Впервые митоз был описан русским ботаником И.Д. Чистяковым в спорах плаунов в 1874 году.
Митоз включает в себя два цитологических процесса: кариокинез и цитокинез.
Кариокинез – деление ядра, цитокинез (цитотомия) – деление цитоплазмы.
В более узком смысле слова под митозом понимают только кариокинез.
Фазы митоза впервые описал немецкий цитолог Вальтер Флеминг, он же ввел термины «митоз», «амитоз», «кариокинез», «хроматин».
Характеристика цитокинеза.
Животные клетки.
Цитокинез происходит путем перешнуровки ее тела, начиная от периферии по направлению к центру. В конце анафазы, благодаря сокращению актиновых микрофиламентов, концентрирующихся по окружности (сократимое кольцо), начинает образовываться клеточная перетяжка, которая углубляясь в телофазе приводит к цитокинезу. Пока еще не завершилось деление цитоплазмы, клетки в течение некоторого времени остаются суживающимися цитоплазматическим мостиком (срединным тельцем), содержащим пучок микротрубочек. Перешнуровка цитоплазмы завершается формированием двух дочерних диплоидных клеток с идентичным набором хромосом. В телофазе происходит распределение органоидов между дочерними клетками, относительной равномерности этого процесса способствует то, что одни органоиды достаточно многочисленны, а другие все время распадаются на мелкие фрагменты и пузырьки.
В растительной клетке цитокинез происходит путем образования предшественника клеточной стенки (фрагмопласта) от центра к периферии. Главную роль в этом процессе играет аппарат Гольджи. Пузырьки, отделяющиеся от аппарата Гольджи и несущие ферменты и компоненты клеточной стенки скапливаются по экватору клетки. Впоследствии они сливаются, образуя мембрану, а их содержимое вытесняется, и из него строится клеточная стенка.
Биологическое значение митоза
1. Обеспечивает равномерное распределение наследственного материала в дочерние клетки.
2. Способствует сохранению кариотипа вида.
3. Является цитологической основой роста тканей и органов
4. Является цитологической основой бесполого размножения.
Типы митоза
1. Стволовой
После него образуются две одинаковые клетки, которые далее размножаются, давая группу однородных клеток
2. Ассиметричный
После него образуются две разные клетки. Одни способны к нормальному делению, а вторая или не способна, или дает начало клеткам, прекращающим размножение через несколько поколений.
3. Трансформирующий
Образовавшиеся после него дочерние клетки претерпевают необратимые изменения. Например, в кожном эпителии клетки, перемещающиеся из базального слоя, накапливают в цитоплазме роговое вещество и утрачивают способность к делению.
Регуляция клеточного цикла.
Осуществляется сложной сбалансированной системой механизмов, стимулирующих и ингибирующих клеточное деление.
Система регуляции клеточного цикла получает и перерабатывает 2 вида информации:
1) информация о действии на клетку различных внешних факторов, стимулирующих или тормозящих ее деление. Клетка перерабатывает эту информацию и определяет свой дальнейший путь (вступление в митоз или в период G0).
2) о инактивности генома.
При повреждении генома клетки прохождение его цикла останавливается и включается система репарации ДНК. Сигналы о повреждении передаются на ген р53, который блокирует прохождение клеткой митотического цикла до устранения повреждения. Если повреждение слишком серьезно, то этот ген запускает программу апоптоза (запрограммированной гибели клетки). Регуляция клеточного цикла возможна, благодаря взаимодействию внутриклеточных факторов: белков – активаторов S-периода, митоз-стимулирующих и митоз-задерживающих факторов, белков циклинов. S-стимулирующие факторы обеспечивают переход клетки из G1 в S. М-стимулирующие белки обеспечивают вступление клетки в митоз, а М-задерживающие факторы тормозят этот процесс. Действие М-стимулирующих факторов проявляется только в присутствии белков циклинов, которые синтезируются на протяжении всего митотического циклы и распадаются в середине митоза.
Общий контроль активности деления клеток осуществляют протоонкогены и антионкогены, а также ряд белковых факторов.
Протоонкогены – группа генов-активаторов, контролирующих нормальное клеточное деление и дифференцировку. Они обуславливают выработку особых белков, которые регулируют деление клеток. Известно более 50 онкогенов. Усиление их активности вызывает развитие опухоли.
Антионкогены – гены, продукты которых угнетают митотическую активность. Например, р53. Продукт этого гена, белок р53 индуцирует синтез продуктов других генов, которые блокируют работу ферментов циклинзависимых киназ, обеспечивающих переход из G1 в S.
Потеря функций гена р53 приводит к утрате контроля над клеточным циклом и способствует развитию опухоли.
Факторы роста:
Это белки-стимуляторы деления клеток, усиливающие М-активность в определенных тканях через специальные рецепторы, присущие определенным клеткам.
Факторы роста:
Это белки-стимуляторы деления клеток, усиливающие М-активность в определенных тканях через специальные рецепторы, присущие определенным клеткам.
Кейлоны – гормоноподобные белки-регуляторы, угнетающие деление клеток. Образуются всеми зрелыми дифференцированными клетками и локально действуют на зрелые клетки той же ткани, способные к делению. Интенсивность клеточных делений определяют с помощью митотического индекса. Это отношение числа клеток, находящихся в митозе к общему числу просмотренных клеток, выраженное в промилях, т.е. в числе митозов на 1000 клеток тканей.
Классификация клеток по уровню их обновления
1. Стабильные клеточные популяции
Состоят из клеток с полной потерей способности к делению. Число клеток в такой популяции стабилизируется в начале их дифференцировки. По мере старения происходит их естественная убыль без восполнения количества
2. Растущие клеточные популяции
Способны к увеличению массы ткани за счет клеточных делений. Это долго живущие клетки, которые выполняют специализированные функции, но сохраняют способность при стимуляции вновь вступать в митотический цикл восстановления нормальной численности (клетки почек, печени, поджелудочной и щитовидной желез, паренхимной ткани).
3. Обновляющиеся клеточные популяции
Характеризуются постоянным обновлением постоянно живущих клеток (клетки эпителия, кишечника, крови, корневого чехлика).
Нарушения нормального хода митоза
Правильное течение митоза может нарушаться различными внешними воздействиями (химическими веществами, некоторыми видами излучений). Такие внешние воздействия могут приводить к нарушению структуры хромосом и даже к изменению их количества при формировании новых дочерних клеток. Например, если произойдет отрыв центромерного района хромосомы, то образуется бесцентромерная хромосома. Такая хромосома не сможет прикрепляться к микротрубочке веретена деления, а следовательно и передвигаться в ходе митоза.
Если перед осуществлением митоза такая хромосома располагается у одного полюса клетки, то она попадет в ядро той клетки, которая образуется у этого полюса. После того, как произойдет расщепление хромосом на сестринские хроматиды, в данной клетке окажется на одну хромосому больше, чем в нормальном кариотипе (2n+1), а вторая клетка, которая сформировалась у противоположного полюса не получит бесцентромерную хромосому и ее кариотип неполным (2n-1).
Если бесцентромерная хромосома в профазе находилась близко к экватору, то вероятнее всего в силу невозможности передвижения в прометафазе она останется на месте и не включится ни в одно из новых ядер. Тогда обе дочерние клетки будут иметь набор хромосом 2n-1.
Некоторые химические вещества (спирты, эфиры) нарушают согласованность процессов митоза. Одни хромосомы начинают двигаться быстрее, а другие отставать, и отставшие хромосомы могут не успеть включиться в формирующееся ядро. Иногда во время деления образуется более двух полюсов. Тогда нарушится механизм распределения хромосом.
Метафазная хромосома двухроматидна, поэтому в анафазе она может взаимодействовать одновременно только с двумя полюсами. Если же полюсов больше, то выбор взаимодействия с определенными полюсами происходит случайно и в результате дочерняя клетка получает не весь набор хромосом. Клетки, получившие неполный хромосомный набор, чаще всего погибают.
Эндорепродукция.
Репликация хромосом без деления ядра. Причиной эндорепродукции может быть завершение митоза из-за блокады определенного этапа митотического цикла. Такая блокада может наступить при переходе из G2 в М, в профазе или метафазе при нарушении цитокинеза. В таких случаях наблюдается явление полиплоидии, т.е. увеличение хромосомного набора ядра клетки. Причина появления полиплоидных ядер заключается в том, что в клетке происходит репликация ДНК, а последующее деление ядра отсутствует.
К особым случаям эндорепродукции относятся политения, эндомитоз, К-митоз.
Политения – репликация хромосом без увеличения их числа за счет того, что реплицированные хроматиды остаются лежать плотно примыкая друг к другу. После репликации ДНК новые дочерние хромосомы продолжают оставаться в деспирализованном состоянии, располагаются друг около друга, не расходятся и в дальнейшем не претерпевают митотическую спирализацию и конденсацию. При многократных репликациях и нерасхождениях хромосомных нитей образуются политемные (гигантские) хромосомы. Они являются интерфазными и никогда не вступают в митоз. По объему они в 1000 раз больше митотических. Политемные хромосомы встречаются в клетках слюнных желез, кишечника, мальпигиевых сосудов, жирового тела, личинок некоторых двукрылых насекомых (в частности дрозофил), в макронуклеусе некоторых инфузорий, в синергидах, некоторых луковых, в некоторых антиподах. Политемные хромосомы неоднородны по своей длине. Они состоят из дисков, междисковых участков и пуфов. Диски – участки конденсированного хроматина, междисковые участки – участки более рыхлых хроматиновых нитей, пуфы – вздутия на местах некоторых дисков, возникающие за счет их деконденсации и разрыхления. Это места транскрипции, здесь синтезируются РНК.
Эндомитоз – увеличение числа хромосом за счет их репликации без деления ядра клетки. Происходит эндомитоз в результате нарушений веретена деления. В случае эндомитоза наступает спирализация хромосом, но ядерная оболочка не исчезает. Так как веретено деления не функционирует, то невозможно расхождение сестринских хроматид к полюсам клетки. Обе сестринские хроматиды каждой хромосомы остаются в одном ядре и в итоге образуются полиплоидные клетки. В эндомитозе различают эндопрофазу, эндометафазу, эндоанафазу и эндотелофазу. Эндомитоз имеет функциональное значение, при нем не прерывается деятельность. Полиплоидные ядра, образованные в эндомитозе, могут вступать в митоз.
Эндомитоз можно вызвать искусственно специальными веществами – цитостатинами, препятствующими образованию веретена деления. Наиболее сильным из цитостатинов является колхицин – это алкалоид – растительный яд, добываемый из растения семейства Лилейные Безвременника осеннего. Эндомитозы, вызванные колхицином, называют К-митозом. Колхицин разрушает микротрубочки веретена деления и препятствует их сборке. К-митозы используют для получения полиплоидных форм растений и при отдаленной гибридизации.
Особый процесс появления полиплоидных соматических клеток происходит у млекопитающих. После S-периода клетки с количеством ДНК 4с вступают в митоз, проходят все его стадии, приступают к цитокинезу. Образуется дочерняя клетка (2n+2n). Она снова вступает в S-период, после чего будет иметь 4с ДНК и 4n хромосом. Затем происходит митоз и последующее объединение двух ядер с образованием одного ядра с набором 8n, а затем нормальное деление с образованием тетраплоидных клеток. Таким образом образуются клетки печени, эпителия мочевого пузыря и некоторые другие.
Результатом эндорепродукции является увеличение размеров клеток и повышением их продуктивности.