В настоящее время взгляды ученых о природе биоэлектрических явлений пользуются всеобщим признанием. Согласно их представлениям, наличие электрических потенциалов в живых клетках обусловлено различной концентрацией ионов Nа+, К+, Са2+ и Сl- внутри и вне клетки, а также различной проницаемостью для них клеточной мембраны. Различают 2 вида биоэлектрических явлений: мембранный потенциал покоя и потенциал действия.
Природу возникновения мембранного потенциала объясняет мембранно-ионная теория, которую предложил Ю. Беpнштейн, а модифицировали А. Ходжкин, Э. Хаксли, Д. Экклс, Б. Катц.

.....…причина возникновения разности потенциалов - неравномерное распределение ионов по обе стороны клеточной мембраны....
Следует отметить, что когда Э. Дюбуа-Реймон выдвинул свою гипотезу в 1846 г, она просуществовала почти 25 лет, а затем в ней усомнились его ученики - Лудимар Герман и Юлиус Бернштейн. Герман считал, что никаких потенциалов и токов в целой неповрежденной мышце не существует. Бернштейн же утверждал, что потенциалы существуют как поврежденной, так и в целой мышце. И именно он победил и заложил основы мембранной теории биопотенциалов.
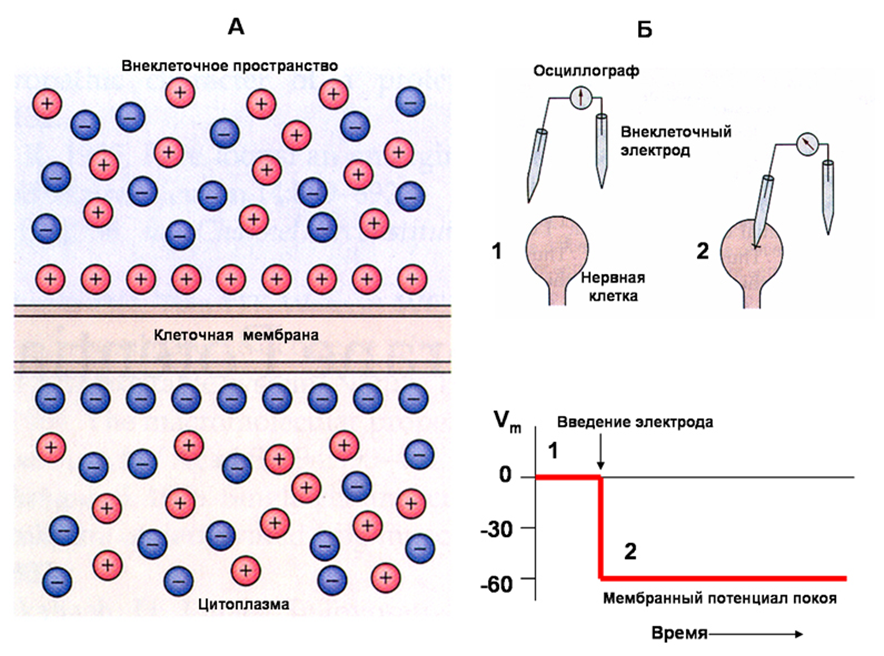
Мембранный потенциал измеряют внутриклеточным электродом, принимая за 0 внеклеточную среду. Поэтому, говоря о мембранном потенциале имеют ввиду внутриклеточный заряд. Внутренняя сторона мембраны по отношению к наружной заряжена отрицательно, поэтому и мембранный потенциал имеет знак «–». В возбудимых тканях мембранный потенциал называют потенциал покоя. Он присущ возбудимым клеткам (нервным, мышечным и секреторным), находящимся в покое, то есть в невозбужденном состоянии.
Потенциал покоя возникает в результате двух причин:
1) Неодинакового распределения ионов по обе стороны мембраны. Внутри клетки находится больше всего ионов К+, снаружи его мало. Ионов Nа+ и ионов Сl- больше снаружи, чем внутри. Такое распределение ионов называется ионной асимметрией.
Благодаря такому распределению ионов, клетки млекопитающих окружены раствором, похожим на разбавленную морскую воду, что является свидетельством пути их эволюции.
2) Избирательной проницаемости мембраны для ионов. В состоянии покоя мембрана неодинаково проницаема для различных ионов. Клеточная мембрана хорошо проницаема для ионов К+, малопроницаема для ионов Nа+ и непроницаема для органических веществ.
За счет этих двух факторов создаются условия для движения ионов. Это движение осуществляется без затрат энергии путем пассивного транспорта — диффузии в результате разности концентрации ионов.
Благодаря наличию на мембранах возбудимых клеток каналов утечки, в частности, для ионов калия, К+ выходит из клетки и увеличивает положительный заряд на наружной поверхности мембраны. Напротив, некоторые открытые каналы для ионов хлора пассивно пропускают Cl- внутрь клетки, что приводит к увеличению положительного заряда на наружной поверхности клетки. Ионы натрия из-за плохой для них проницаемости мембраны в состоянии покоя, накапливаются на наружной поверхности мембраны и увеличивают ее положительный заряд. Органические соединения (например, белки, фосфаты), имеющие на поверхности отрицательный заряд, остаются внутри клетки.
В результате такого движения наружная поверхность мембраны заряжается положительно, а внутренняя - отрицательно. Внутренняя поверхность мембраны может не быть абсолютно отрицательно заряженной, но она всегда заряжена отрицательно по отношению к внешней. Такое состояние клеточной мембраны называется состоянием поляризации. Движение ионов продолжается до тех пор, пока не уравновесится разность потенциалов на мембране, т. е. не наступит электрохимическое равновесие.
В возникновении мембранного потенциала участвует сила диффузии (разность концентрации ионов) и сила электростатического взаимодействия, поэтому мембранный потенциал имеет концентрационно-электрохимическое происхождение.
Для поддержания ионной асимметрии электрохимического равновесия недостаточно. В клетке имеется другой механизм - натрий-калиевый насос, обеспечивающий активный транспорт ионов.
Работа натрий-калиевого насоса обеспечивает 1) высокую концентрацию ионов К+ внутри клетки, т.е. постоянную величину потенциала покоя; 2) низкую концентрацию ионов Nа+ внутри клетки, т. е. сохраняет нормальную осмолярность и объем клетки, создает базу для генерации потенциала действия.
Таким образом, мембранный потенциал покоя представляет собой разность потенциалов между наружной и внутренней поверхностью мембраны клетки.
Он является результатом разделения зарядов относительно клеточной мембраны, которое возникает за счёт движения заряженных ионов (натрия и калия) по концентрационным градиентам через ионные каналы, открывающиеся в покое.
Мембранный потенциал в состоянии покоя клетки имеет отрицательный знак (значение).
Наличие потенциала на мембране возбудимой клетки лежит в основе механизмов возникновения в ней электрических распространяющихся сигналов – потенциалов действия.