
Род Streptococcus включает в себя основные бактерии, вызывающие мастит у коров. Мастит — это распространённое и дорогостоящее заболевание в молочной промышленности, которое приводит к потерям молока и затратам на лечение, а также негативно сказывается на здоровье животных. В этом обзоре рассматриваются виды Streptococcus, наиболее часто вызывающие мастит у коров (S. agalactiae, S. dysgalactiae и S. uberis), их классификация, источники и частота возникновения.
Обсуждаются факторы, связанные с животными и окружающей средой, которые влияют на восприимчивость к маститу и уровень заражения, поскольку было установлено, что Streptococcus широко распространён в стаде, но не всегда приводит к маститу. Также рассматриваются иммунные реакции коров после заражения и различия в вирулентности основных видов Streptococcus.
Описаны различные методы выявления мастита и возможные варианты лечения, включая традиционные и альтернативные методы. Основным способом лечения стрептококкового мастита являются ß-лактамные антибиотики, однако у Streptococcus выявлена устойчивость к некоторым другим антибиотикам, таким как энрофлоксацин, тетрациклин и эритромицин.
В конце рассматриваются меры контроля и профилактики, включая вакцинацию, гигиену и другие вмешательства. Цель обзора — предоставить подробную информацию о роли Streptococcus в развитии мастита у коров.
Введение
Существует более 130 патогенов, связанных с маститом у коров, и многие из них относятся к роду Streptococcus. Эти стрептококки — грамположительные бактерии сферической формы (0,5–2 мкм), которые обычно образуют пары или цепочки. Их классификация основана на морфологии колоний, типе гемолиза и серологической специфичности в системе Лэнсфилда. Многие стрептококки являются факультативными анаэробами и непатогенны, являясь частью нормальной микробиоты человека и животных. Однако некоторые виды могут вызывать серьёзные заболевания, включая мастит у крупного рогатого скота. Среди них наиболее значимы S. agalactiae, S. dysgalactiae и S. uberis. Редко встречающиеся патогены, вызывающие мастит, включают S. canis, S. lutetiensis и S. equinus.
Мастит является самым распространённым и дорогостоящим заболеванием в молочной промышленности во всем мире. Экономические потери от мастита составляют около 124 евро на корову в год, что приводит к потерям в 500 миллионов евро в Германии, 3 миллиарда евро в ЕС и 125 миллиардов евро по всему миру. Мировая распространённость мастита у коров составляет от 30 до 50% всех коров в год. Помимо финансовых потерь, связанных с уменьшением надоев, снижением качества молока, затратами на ветеринарное лечение и лекарства, мастит представляет серьёзную проблему для здоровья и благополучия животных и является одной из главных причин выбраковки молочных коров.
Симптомы мастита включают отёк, жар и боль в вымени, ненормальный вид молока, повышенную температуру тела, вялость и анорексию. Мастит можно разделить на три категории в зависимости от степени воспаления: клинический, субклинический и хронический. Клинический мастит характеризуется явными отклонениями в состоянии коровы и молока, тогда как при субклиническом мастите изменения касаются только надоев и количества соматических клеток в молоке. Частота субклинического мастита в 15–40 раз выше, чем клинического, что делает его более значимым с экономической точки зрения из-за его способности снижать надои, оставаясь при этом незамеченным.
Средняя продолжительность стрептококкового мастита составляет 12 дней, но в хронических случаях он может длиться более 300 дней. Нелеченый острый мастит может перейти в хронический и привести к снижению фертильности. S. uberis и S. agalactiae известны как патогены, способные вызывать хронический мастит. Лечение и профилактика мастита — основные причины использования антибиотиков у молочных коров, что увеличивает риск развития устойчивости микроорганизмов к противомикробным препаратам.
Классификация
Стрептококки являются одними из ключевых патогенов, вызывающих мастит у крупного рогатого скота по всему миру. Эти патогены можно разделить на две основные группы: инфекционные и экологические.
Инфекционные патогены приспособлены к выживанию внутри хозяина и передаются от коровы к корове, в основном через доение. Эти бактерии могут быстро и широко распространяться в стаде.
Экологические патогены, наоборот, могут выживать вне хозяина и часто являются частью нормальной микрофлоры, окружающей корову. Заражение ими может происходить во время и между доениями, в период сухостоя или перед отелом. Влияние экологических факторов, таких как влажность и температура, существенно влияет на распространение этих патогенов.
S. uberis обычно классифицируется как экологический патоген, хотя зарегистрированы случаи передачи между коровами. Этот вид может быть как альфа-гемолитическим, так и негемолитическим. Он характеризуется переменным фенотипом в тесте CAMP, а также способностью расщеплять эскулин, гиппуровую кислоту и инулин. S. uberis обнаруживается у различных видов крупного рогатого скота, включая дойных и сухостойных коров, телок и многоплодных коров.
S. dysgalactiae имеет смешанную классификацию, так как способен выживать как внутри хозяина, так и в окружающей среде, и часто описывается как промежуточный патоген. Этот вид обычно негемолитический и классифицируется как группа C по системе Лэнсфилда. S. dysgalactiae чаще всего ассоциируется с инфекциями крупного рогатого скота, но может также поражать других жвачных животных, таких как козы и овцы.
S. agalactiae является инфекционным патогеном, способным колонизировать желудочно-кишечный тракт молочных коров. Он может долгое время выживать в молочных железах, образуя биопленки, и часто связан с субклиническим маститом. Этот вид входит в группу B по классификации Лэнсфилда и обычно считается бета-гемолитическим, хотя существуют негемолитические штаммы. S. agalactiae имеет множество серотипов, различающихся по патогенности и влиянию на человека и животных.
S. canis является инфекционным патогеном из группы Lancefield G, характеризуется бета-гемолитическим, CAMP-отрицательным и аэскулин-гидролизующим фенотипом.
S. equinus классифицируется как экологический патоген из группы Lancefield D, его фенотип характеризуется изменчивым гемолизом, он CAMP-отрицателен и гидролизует аэскулин.
S. lutetiensis также относится к инфекционным патогенам, входит в группу Lancefield D, характеризуется альфа-гемолизом и CAMP-отрицательным, а также гидролизом аэскулина.
Резервуары, возникновение и частота стрептококкового мастита
Как уже упоминалось, S. uberis обитает в основном в среде обитания коров и является наиболее распространённым стрептококком, вызывающим мастит, с растущей распространенностью по всему миру. Чаще всего его находят в подстилочном материале, особенно в соломе, где его количество намного выше, чем в песке или опилках. Места для лежания коров и их подстилки являются основными точками проникновения S. uberis в вымя, так как они характеризуются высокой бактериальной нагрузкой. Также существует близкородственный вид — S. parauberis, который фенотипически неотличим от S. uberis, но отличается продукцией β-d-глюкуронидазы. Однако роль S. parauberis в развитии маститных инфекций незначительна из-за её очень низкой распространенности, составляющей примерно 0,5% всех случаев мастита.
Для S. dysgalactiae значительную роль в возникновении и поддержании маститных инфекций играет обычная муха для крупного рогатого скота — Hydrotaea irritans.
S. agalactiae в основном встречается в вымени коровы, но недавние исследования показали, что желудочно-кишечный тракт коров и окружающая среда (полы, подстилка и питьевая вода) также могут быть резервуарами этой бактерии.
S. canis, S. equinus и S. lutetiensis редко ассоциируются с маститом крупного рогатого скота и были диагностированы менее чем в 1% всех стрептококковых инфекций. Резервуары и факторы риска для этих видов стрептококков в значительной степени неизвестны. Мастит, вызванный S. canis, в основном передаётся горизонтально от кошек и собак, живущих рядом с коровами.
Ранее заразные патогены были гораздо более проблематичными. В 1927 году S. agalactiae был ответственен за 90% всех случаев мастита. Однако с введением пятипунктного гигиенического плана в 1960-х годах частота мастита, вызванного заразными патогенами, значительно снизилась. План включал быстрое выявление и лечение инфицированных коров, антибиотикотерапию сухостойных коров, дезинфекцию сосков после доения, убой хронически больных коров и регулярную дезинфекцию доильного аппарата. Эти меры привели к значительному снижению общего числа случаев мастита (на 75%), особенно для заразных патогенов, таких как S. agalactiae. В настоящее время большинство случаев мастита (примерно 90%) вызваны патогенами окружающей среды, причем доля S. uberis составляет более одной трети всех случаев в среднем по миру.
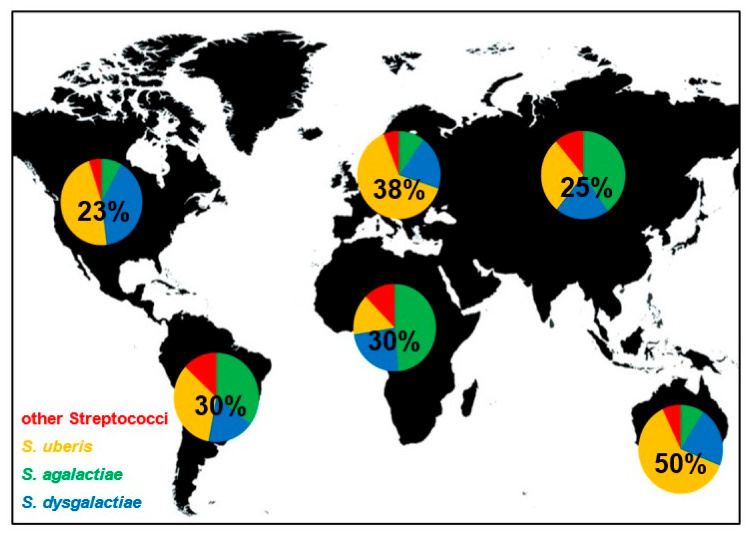
Хотя контагиозные и экологические стрептококки вызывают мастит у крупного рогатого скота по всему миру, их распределение неравномерно. В Европе и Австралии/Новой Зеландии S. uberis вызывает большинство случаев стрептококкового мастита (64% и 62% соответственно). В Африке и Азии S. agalactiae является основным возбудителем (49% и 40% соответственно). В Северной Америке доминирующими патогенами являются S. uberis и S. dysgalactiae с аналогичной частотой заболеваемости (47% и 40% соответственно). В Южной Америке наибольшие и схожие частоты мастита вызваны S. uberis и S. agalactiae (34% и 35%).
Частота стрептококкового мастита также варьируется по регионам. Наиболее высокая частота наблюдается в Австралии (50%), затем следует Европа (38%). На других континентах стрептококковый мастит встречается реже и ответственен за 23–30% всех инфекций мастита. Эти данные предоставляют широкий обзор распространения стрептококкового мастита по всему миру и показывают различия в распространенности видов в разных регионах.
Эпидемиологические факторы, влияющие на восприимчивость к стрептококковому маститу
Разница в распространенности стрептококков и количестве случаев мастита среди коров в разных стадах обусловлена различными факторами. Влияние на восприимчивость коров к стрептококковому маститу оказывают как характеристики самих коров, так и условия их содержания.
На восприимчивость к маститу влияют:
- Возраст коровы: У старых коров (старше 10 лет) мастит встречается на 24% чаще, чем у молодых (3-6 лет). Это связано с тем, что у старых коров сосковый канал шире или всегда частично открыт из-за частого доения.
- Стадия лактации: Коровы на ранней стадии лактации (0-3 месяца) подвержены более высокому риску мастита. В середине лактации (4-6 месяцев) риск самый низкий, а на поздней стадии (более 7 месяцев) риск снова увеличивается.
- Количество родов: Чем больше родов у коровы, тем выше риск мастита. У коров с более чем шестью отелами вероятность заболеть маститом на 30% выше, чем у коров с 1-3 отелами.
- Гигиеническая практика доения: Хорошая гигиена, например, мытье вымени до и после доения, существенно снижает риск мастита.
Эти факторы помогают объяснить, почему частота стрептококкового мастита варьируется среди коров в разных стадах и даже в пределах одного стада.
Во время лактации корове требуется больше энергии и питательных веществ. Питание влияет на иммунитет коровы и может быть связано с частотой мастита. Концентрация витамина Е и селена в крови и тканях у коров самая низкая в период отела. Добавление селена и витамина Е в корм помогает снизить частоту стрептококкового мастита.
Восприимчивость коров к маститу также определяется рядом генетических признаков. Установлена положительная корреляция между возникновением мастита и удоями молока, что означает, что селекция на более высокие удои приводит к увеличению числа случаев мастита. Программы селекции, направленные на повышение устойчивости к маститу, в основном акцентируют внимание на таких характеристиках, как низкий уровень заболеваемости маститом, низкое содержание соматических клеток (SCC) в молоке, утечка молока и скорость доения. SCC считается наиболее полезным косвенным показателем устойчивости к маститу, поскольку между ними существует высокая генетическая корреляция, достигающая 60–85%. Длина соска, его форма и время закрытия после доения также различаются у коров и являются важными генетическими признаками, влияющими на восприимчивость к маститу. Действительно, до 70% сосков могут быть загрязнены экологическими стрептококками до доения, а кончик соска является основной точкой проникновения патогенов. Таким образом, риск мастита значительно зависит от длины соска, его формы и времени полного закрытия после доения и в период сухостоя.
Состав микробиоты вымени и история мастита также коррелируют и играют важную роль в определении восприимчивости к маститу. Микробиота вымени здоровых коров значительно отличается от микробиоты коров, перенесших мастит. Здоровые четверти вымени демонстрируют более высокое таксономическое разнообразие по сравнению с четвертями, имеющими в анамнезе мастит. Данные свидетельствуют о том, что микробиота изменяется в вымени после перенесенного мастита, даже если инфекция произошла давно. Неоптимальное разнообразие микробиоты в вымени, ранее пораженном маститом, может объяснять повышенную восприимчивость коров к повторным случаям мастита.
В дополнение к ранее упомянутым факторам, влияющим на восприимчивость коров к маститу, на показатели мастита также оказывают влияние факторы окружающей среды, такие как время года и методы управления стадом. В частности, количество случаев контагиозного стрептококкового мастита тесно связано с гигиеническими условиями доения. Показатели мастита значительно снижаются (-64%), когда вымя моют до и после доения, по сравнению с ситуацией, когда очистка не проводится. Наиболее высокие показатели мастита наблюдаются летом, что может быть связано с высокими температурами и, соответственно, с повышенной скоростью распространения возбудителей мастита в окружающей среде или передачей бактерий мухами, особенно активными в это время года. Известно, что бактерия Streptococcus dysgalactiae, связанная с летним маститом, переносится мухами, особенно в Северной Европе и Японии.
Факторы патогенеза и вирулентности
В ходе инфекционного процесса стрептококки сталкиваются с разнообразными меняющимися условиями и, следовательно, должны адаптироваться к различным условиям для успешного инфицирования. Как следствие, они экспрессируют ряд факторов вирулентности, которые позволяют выживать и реплицироваться в различных тканях хозяина во время патогенеза, например, путем придания адгезии и инвазии клеткам хозяина или избегания иммунного распознавания и последующей деградации. Факторы вирулентности S. uberis, S. dysgalactiae и S. agalactiae суммированы.
Среди всех перечисленных факторов вирулентности исследование 78 штаммов S. uberis показало, что три наиболее распространенных — это ген hasC (продукция гиалуроновой кислоты), присутствующий в 89,7% исследованных штаммов, за которым следует ген sua (связывание лактоферрина) в 83,3% и gapC (связывание плазмина, а также иммуномодуляция) в 79,4%. Оперон has — это консервативная область гена, присутствующая во всех штаммах стрептококков группы A и инкапсулированных стрептококках группы C. Он обеспечивает образование капсулы из гиалуроновой кислоты, которая защищает, например, S. uberis от опсонизации и фагоцитоза и опосредует устойчивость к бактериальному очищению внутри нейтрофильных внеклеточных ловушек.