Я студентам люблю рассказывать про так называемые контуры разрушения. В норме биогеоценоз[1] представляет собой гомеостат.
В ответ на разнообразные нарушения — рубки, гари, потравы от рекреации и прочее — включается отрицательная обратная связь в виде восстановительных сукцессий (демутаций). Они восстанавливают исходный живой покров с присущей ему мозаичностью, условиями питания, почв и влажности и прочее, но — только пока «следов» внешних воздействий в виде разных типов нарушений немного.
Когда же их «сеть» переходит некий предел плотности и/или интенсивности нарушений[2], вместо ожидаемой отрицательной обратной связи запускается положительная, идут изменения, внешне сходные с восстановительными сукцессиями, однако не ликвидирующие нарушения, а, напротив, усиливающие их: распространяющие в пространстве (их отдельные «пятна» ползут и сливаются между собой, как прореха на гнилой ткани) и усиливающие нарушенность в каждом отдельном «пятне». В пределе это ликвидирует прежнее сообщество и устанавливает на месте его новое, антропогенно изменённое.
Дальше рассказывается о трёх самых ярких и «свежих» примерах этого. Первый — явственный крах надежд на «глобальное позеленение» (англ. global greening): планетарный прогресс растительности, увеличение фитомассы, восстановление и экспансия природных биомов, реагирующих на «удобрение углекислым газом» — экспоненциальный рост содержания его в атмосфере в последние 50 или 100 лет вследствие антропогенных причин, ответственный за глобальное потепление. В последние 20-25 лет надёжно показан противоположный процесс — рост «сжигания» углерода, «запасённого» в торфах, мортмассе, углероде почвы по ходу глобального потепления вместо чаемого увеличения его связывания растительностью или захоронения болотами, именно потому что биомы планеты уже 50 (или 100) лет как уж слишком нарушены.
Рост «сжигания» запасённого углерода вместо «удобрения углекислым газом»
После того как в 2015 г. было надёжно показано, что глобальное потепление есть, и причина его — деятельность человека, отрицатели того и другого сменили пластинку, стали рассказывать, что «от потепления будет только лучше»: подъём содержания углекислого газа, идущий неукротимо, стимулирует развитие растительности, экспансию лесов, зарастание пустынь, рост благоприятности климата вместо роста экстремальности, ожидаемой алармистами, в общем, и на Марсе будут яблони цвести.
Эта гипотеза «глобального озеленения» впервые высказывалась в 2000-х и сильно набрала популярность в последние годы.
Известно, что в настоящей качественной дезинформации должно быть около 60% правды, если для публики, или 95%, если для специалистов.
«Глобальное озеленение» ближе ко второй цифре, в ней есть рациональное зерно, важное и интересное нам, биологам: это гипотеза «противодействия изменению», отражающая способность биосферы регулировать возмущение в виде роста выбросов CO₂ и СН₄, как то было в доагрикультурный период (Фридман, Ерёмкин, 2015).
Но, взятая вне контекста реальных биомов планеты[3], разнотипных и, увы, более чем на трети[4] исходной площади уже ликвидированных (либо сильно нарушенных) человеком, эта верная мысль делается ошибкой или дезинформацией (в зависимости от упорства, с которым на ней настаивают вопреки множеству не ложащихся в данное объяснение фактов, причём разнотипных). В отличие от ситуации в плиоцене, когда сравнимая концентрация парниковых газов имела сугубо положительные последствия (Сахара по большей части была саванной, там текли реки и прочее), сегодняшний экспоненциальный рост концентраций главных парниковых газов, CO₂ и СН₄ — отнюдь не отдельный природный процесс, но интегральная часть антропогенного разрушения природных биомов и его следствие[5].
Важнее всего, что «глобальное озеленение» напрочь опровергается фактом, как раз относящимся к «контурам разрушения» (есть и другие, но это отдельная тема). Вместо ожидаемого запуска отрицательной обратной связи (усиления фотосинтетического связывания CO₂ и захоронения С в виде торфа, выводящего углерод из круговорота — главный путь «охлаждения» планеты биотическими процессами) в нарушенной биосфере, увы, реализуется связь положительная. Рост концентрации CO₂ ещё сильней ускоряет «проедание» органики, ранее захоронённой в торфах и других долговременных «хранилищах» углерода: опаде, мёртвой древесине, подстилке и почве (её объём показан в рис.1). А это, в свою очередь, ускоряет рост концентрации CO₂ в атмофере: круг замыкается.
Рисунок 1. Запасы углерода в лесах разных климатических поясов, 2000e гг. (а) и в 14 главнейших биомах планеты, более подробные оценки 2010х гг. (б).
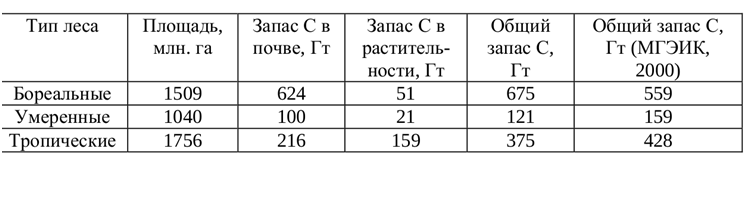
Оказывается, что экосистемы поглощают всё меньшую долю от огромного количества углекислого газа, ежегодно попадающего в атмосферу из-за сжигания ископаемого топлива, производства цемента, осушения болот, выжиганий растительности, других нарушений экосистем. До недавнего времени рост выбросов CO₂ в атмосферу сопровождался и большим связыванием его растениями в ходе фотосинтеза, в меньшей степени и фитопланктоном океана. Однако с 2000-х гг. рост связывания CO₂ и захоронения С уже «не поспевал» за ростом его содержания в атмосфере, с 2010-х он и вовсе стал падать.
Впервые это выявили при изучении межгодового сдвига сезонных колебаний концентраций CO₂ в Северном полушарии[6]. По мере развития глобального потепления вёсны приходят всё раньше, раньше начинается фотосинтез, поэтому связывание CO₂ вроде бы больше и больше должно преобладать над дыханием. Однако результат опроверг ожидания: изменения климата в экосистемах сильней стимулировали дыхание, нежели фотосинтез, особенно осенью, так что по мере прогресса потепления даты осени, когда дыхание начинает преобладать над фотосинтезом, с каждым годом приходят всё раньше. Аномально тёплые осени, происходящие всё чаще и чаще, резко усиливают дыхание всех организмов (не только растений, но прежде всего бактерий с грибами) согласно правилу Вант-Гоффа. И это чем дальше, тем больше укорачивает ежегодный сезон преобладания связывания CO₂ над его выделением (вместо чаемого удлинения), чем сводят на нет «прибавку» в связывании С, созданную более ранними вёснами.
« …То, что «углеродная ёмкость» океана уже достигла предела и связывание океаном дополнительного количества CO₂ сокращается, недавно доказано прямыми наблюдениями (Le Quéré C. et al., 2007). … в Nature от 3 января 2008 года опубликована статья, в которой сообщается о тревожных признаках сокращения возможностей поглощения CO₂ также и наземными экосистемами. Авторы … (всего 16 человек) сосредоточили своё внимание на соотношении многолетнего тренда увеличения CO₂ в атмосфере и регулярных сезонных колебаний концентрации CO₂, происходящих на фоне этого тренда.Как эти две динамики (многолетняя и сезонная) соотносятся, можно пояснить на примере самого длинного (почти за полвека) ряда данных, полученных обсерваторией Мауна-Лоа на острове Гавайи. График, суммирующий результаты измерений, показывает как непрерывный рост, так и небольшие, но регулярные сезонные колебания концентрации CO₂: максимум приходится на апрель-май, а минимум на сентябрь-октябрь. Возникают эти колебания из-за того, что процесс потребления углекислого газа, а именно фотосинтез растений, происходит только в тёплый период с конца весны и до окончания лета (в северном полушарии это май-август). Осенью, зимой и в начале весны фотосинтез невозможен (по крайней мере в умеренных и северных широтах, где устанавливаются отрицательные температуры).Но параллельно в экосистеме протекает процесс, обратный фотосинтезу, — дыхание (разложение органического вещества с потреблением кислорода и выделением CO₂). Хотя дышат все организмы, поступление в атмосферу основной массы углекислого газа почти целиком определяется дыханием редуцентов, в основном бактерий и грибов. Дыхание происходит в течение более длительного периода, чем фотосинтез. Летом, когда тепло, интенсивность его особенно велика, но на это же время приходится пик фотосинтеза, и в результате связывается CO2 гораздо больше, чем выделяется. Но как только фотосинтез ослабляется, пропорция потребления и выделения CO₂ сдвигается в сторону выделения, увеличивая концентрацию CO₂ в воздухе».
Авторы обсуждаемой работы на примере нескольких непрерывных (продолжающихся по меньшей мере 15 лет) рядов наблюдений за изменениями содержания CO₂ в разных точках Северного полушария проследили, как меняется во времени положение тех точек на графике, где линия сезонных колебаний пересекает линию основного тренда. За год таких точек две: «весеннее пересечение» соответствует моменту, когда кривая содержания CO₂ идёт вниз: в результате интенсивного фотосинтеза процессы связывания этого газа начинают преобладать над выделением. «Осеннее пересечение» соответствует моменту, когда кривая идёт вверх, и выделение CO₂ в результате дыхания преобладает над связыванием (рис.2б).
Доселе предполагалось, что увеличение потребления CO₂ растительностью происходит прежде всего за счёт удлинения сезона вегетации — периода активного роста растений. Действительно, весна фенологически приходит всё раньше (в Западной Европе по сравнению с 1960 гг. в среднем уже на 12 дней раньше), а осень всё чаще аномально тёплая. По идее, данные изменения климата (и следующий за ним сдвиг фенодат) скажутся и на сезонной динамике содержания CO₂. «Весеннее пересечение» должно наступать всё раньше, а «осеннее» позже (рис. 2б).
На деле же вышло обратное: если «весеннее пересечение» действительно стало наблюдаться раньше, то «осеннее» тоже сдвинулось на более ранние сроки, а не отодвинулось на более поздние вопреки ожиданиям. Благодаря этому резко усилилось выделение CO₂ осенью, перечёркивающее рост связывания CO₂ более тёплой и ранней весны (в 2000е гг. — на 90%, сейчас — полностью и даже с избытком). Piao et al. (2008) показывают, что тогда поглощение углерода северными экосистемами (и не только) заметно сократится в самое ближайшее время, а выделение усилится и, чем дальше, тем больше, будет перекрывать поглощение.
Этот печальный прогноз оправдался с лихвой, ибо данные тренды непросто сохранились, но даже усилились, в 2010-е, распространившись из северных и умеренных широт в тропики (примерно к 2015 г.). Что достигается прежде всего за счёт сдвига «осеннего пересечения» на всё более ранние даты, но не только. Первое зафиксировано раньше всего — в 1990-2000-е гг. Piao et al. (2008) анализировали погодовые изменения дат «осеннего пересечения», аномалий осенней температуры и периода преобладания поглощения углерода (CUP) для 10 станций сети NOAA/ESRL (отбор проб воздуха для определения концентрации CO₂, потоков CO₂ в экосистемах и прочее на протяжении минимум 15 лет непрерывно). Их размещение в Северном полушарии см. красные точки на схеме рис.3б. Также по спутниковым данным оценивали развитие растительности (индекс NDVI[7]), валовую (NEP) и чистую (NPP) продукцию экосистем, а также их дыхание (ТЕР).
За все эти годы «осеннее пересечение» в целом сдвигалось на более ранние даты (рис.3а), они отрицательно коррелируют с аномалиями осенних температур вокруг каждой станции (в широкой полосе ±20O широты), для 4 из 10 мест корреляция значима. Вероятность чисто случайного получения таких результатов 10-3 при независимости записей, производимых на 10 станциях.
Авторы опровергли предположение, что сдвиг «осеннего пересечения» на более ранние даты и негативная корреляция его величины с аномалиями осенней температуры созданы межгодовыми различиями интенсивности ветров, определяющих перенос CO₂ в атмосфере, установив жёсткую связь с погодовыми изменениями валовой продуктивности экосистемы NEP: в тёплые осени её накопление завершается раньше. Они анализировали влияние обоих факторов в LMDZt — модели атмосферной циркуляции; за исключением двух мест наблюдений (гора Симоне и мыс Кумукахи, CMN и KUM) на отклик влияли именно изменения NEP, эффект атмосферного переноса был небольшим. Рост поглощения CO₂ океаном и сжигания углеводородов влиял незначительно, объясняя не более 4% дисперсии даты «осеннего пересечения» (исключая KUM). Изменения сезонности такого сжигания, в связи с климатическими, могут повлиять лишь со временем, и то незначительно. Этот сдвиг не фиксируется в наблюдениях тех же станций за 1970-1994 гг., но сразу возникает потом.
Имитация погодовых изменений продуктивности наземных экосистем Северной Евразии, зависящей от климатических переменных и атмосферного переноса CO₂ (модель ORCHIDEE, «настроенная» по данным 1980-2002 гг.) опровергла[8] конкурирующее объяснение, что увеличение осенних потерь CO₂ и укорочение CUP — следствие большей засушливости лета, предшествующего подобной осени. Слабые летние минимумы поглощения CO₂ (признак засухи) значимо не влияли на данные феномены.
Укорочение CUP, фиксируемое почти на всех станциях, и сохраняющееся в дальнейшем, означает снижение продуктивности всё большего числа экосистем на всё большей площади, сокращение фитомассы вместо её накопления, «сжигание» углерода, ранее «запасённого» экосистемой, вместо ожидаемого увеличения этих запасов. Именно это фиксируется при моделировании отклика потоков углерода через реакцию экосистем на глобальное потепление в ORCHIDEE, см. рис.5, а — для изменений их валовой продуктивности фотосинтеза[9] осенью (GPP, более жёлтый и бурый — снижение, более зелёный и синий — рост, в Тг С/м²*OС), b — для изменений их чистой продукции осенью, c — для изменений их валовой продукции осенью, d — для изменений NDVI.
Этот эффект, созданный «контуром разрушения», в следующие десятилетия не только не ослаб, но резко усилился, а также из северных широт распространился в тропические. Данные по лесам северного полушария показывают сдвиг «весеннего пересечения» вперёд на 1,9±0,1 дня, однако осеннего — уже на 2,6±0,1 дня, то есть CUP сокращается повсеместно (каждый дополнительный день вегетации позволяет поглотить больше углерода, от 3 до 9,8 г/м²). Прямые измерения газообмена на уровне листьев показывают, что это связано с более ранним началом старения и отмирания листьев в тёплые осени (Zohner et al., 2023). Всё это не считая взлёта дыхания редуцентов в тёплые осени!
Вследствие этой и ряда других[10] причин в 1980-2015 гг. фотосинтетический отклик экосистем после короткого подъёма упал, тогда как дыхание постоянно растёт, и к 2040-2060 гг. во всех наземных экосистемах выделение CO₂ превзойдёт поглощение, вместо «глобального позеленения» произойдёт «побурение», global browing. Уже к 2020 г. все биомы планеты на рост содержания CO₂ реагируют ростом его выделения, вместо ожидаемой интенсификации связывания, то есть реализуют классический контур разрушения.
Продолжение следует.
Примечания:
[1] То есть растительность и животный мир, взятые в присущих им почвенных и ландшафтных границах: этим он отличается от понятия «экосистема», включающего только биоту и абстрагирующегося от почвенных, ландшафтных и географических привязок места действия экологических (приспособление) и эволюционных (адаптация) процессов.
[2] Скажем, для разных ландшафтов Приамурья предельно допустимый уровень антропогенной трансформации (после которого исходный ландшафт не восстанавливается даже при снятии нагрузки, а превращается во вторичный) устанавлен Ф.Н. Рянским (1995), см.также таблицу из этой работы.
[3] Разнотипных, включающих не только растительность, но и её потребителей, с «блоком» устойчивости, основанным на обороте мортмассы, когда переменчивость почвенно-климатических условий сдвигает баланс между гумификацией и минерализацией ту или иную сторону, обеспечивая сохранность почвенного плодородия и устойчивость «выдачи» минеральных веществ растениям вопреки названным изменениям. Этот почвенный механизм обеспечивает различия между биомами по устойчивости к стрессам, наряду с различиями в продуктивности и уровне биоразнообразия.
[4] часто до половины и двух третей, в крайних случаях, как европейские степи, и больше.
[5] См. подробней доклад на Ломоносовских чтениях-2023 (каф.высших растений).
[6] Piao Sh., Ciais Ph., Friedlingstein P. et al. Net carbon dioxide losses of northern ecosystems in response to autumn warming // Nature. 2008. V. 451. P. 49-54.
[7] Normalized difference vegetation index, рассчитывается по спутниковым снимкам, показывает количество и развитость растительного покрова (конкретнее плотность фотосинтетически активной биомассы) на данной территории.
[8] Её выводы контролировали тремя независимыми способами: спутниковыми измерениями индекса листовой поверхности (англ. LAI — Leaf area index, мера развития освещённых солнцем фотосинтезирующих поверхностей и, соответственно, продуктивности экосистемы) в те же годы, наземными оценками выделения CO₂ и контролем сезонных колебаний концентрации CO в воздухе.
[9] До потребления возникшего органического вещества разными консументами и редуцентами.
Автор — Владимир Фридман (wolf_kitses), «XX2 ВЕК».
Вам также может быть интересно: