В человеческом организме около 3,72 × 10^3 клеток, а сколько в нас молекул и атомов- даже сложно представить. Удивительно, но это невообразимое число можно свести всего к паре десятков химических элементов. NOCH все наверняка знают: они присутствуют абсолютно во всех органических веществах. Но кроме самых базовых есть элементы, о которых ты вроде слышал, но даже до конца и не уверен, что они есть в твоем организме. Ну да, железо, что-то про кровь, про перенос кислорода. Но у растений, например, нет крови, но железо им все равно нужно, зачем? А если сказать, что оно еще само по себе и токсично, почему же мы все тогда живы, здоровы и счастливы? Ну давайте разбираться.
БАЗА БАЗИРОВАННАЯ
Железо — это d-элемент восьмой группы четвертого периода В живых организмах железо представлено в виде двух ионов: Fe2+ (железо двухвалентное) и Fe3+ (железо трехвалентное).Отдельные молекулы железа потенциально токсичны, поскольку в аэробных условиях они катализируют распространение активных форм кислорода и генерацию высокореактивных радикалов, таких как OH–. Так называемая реакция Фентона: Fe2+ + H2O2= Fe3+ + HO•+ OH-
Fe3++ H2O2= Fe2+ + HOO• + H+
Так что внутри живых организмов они не встречаются в свободном виде и спрятаны в различные структуры для использования, транспорта и хранения.
ЗАЧЕМ ВООБЩЕ НУЖНО ЖЕЛЕЗО?
Если кратко — функционал огромен. Во- первых, то самое что-то про кровь: железо входит во всем известные соединения, связывающие кислород: гемоглобин в эритроцитах и миоглобин в мышцах, на синтез которых и уходит подавляющая часть запасов микроэлемента.
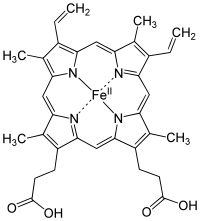
Железо является компонентом большого числа ферментов: каталаз и пероксидаз, и разнообразных по функциям белков- цитохромов. Железо проявляет 2 степени окисления, следовательно, оно может участвовать в окислительно- восстановительных реакциях, что собственно и делает практически во всех указанных соединениях. Именно поэтому железо почти всегда встречается в реакционных центрах белков, а также в входит в состав гемов и Fe-S кластеров, о которых мы поговорим чуть позже.
Две трети железа в организме человека содержится в том самом, известном всем, гемоглобине, железопротеине, который используется для переноса кислорода красными кровяными тельцами — эритроцитами. Эритроцит, как и любая другая клетка человеческого тела, имеет ограниченный срок жизни~111 дней. После гибели клетка поглощается специальными макрофагами в селезенке или печени, а содержащийся в нем гемоглобин «разбирается» обратно, и железо вновь вступает на свой метаболический путь: накапливается в теле или сразу идет в красный костный мозг к предшественникам эритроцитов.
ГЕМЫ И НЕ ГЕМЫ
Железо, присутствующее в организме можно разделить на два вида и обозвать их как гемовое и негемовое. С негемовым железом все просто: это свободные ионы железа, не входящие в органические соединения. В теле животного такое железо аккумулируется в составе запасающего белка печени — ферритина, о котором мы еще будем говорить позже. гемовоежелезо, как легко догадаться, находится в составе гема — сложного соединения с двухвалентным ионом внутри порфиринового кольца.
Порфирин — это тетрапиррол, основная часть гема, окружающая ион железа. В организме гемовое железо входит в состав гемопротеинов (белков, содержащих гем :).
Как мы видим, железо в ходит в состав многих соединений, а специальных систем его вывода из организма не существует. Следовательно, для поддержания постоянной концентрации железа нужно уметь грамотно контролировать его поступление. Чтобы понять, как это происходит, нужно разобраться с тем, как железо вообще попадает в организм.
МЕТАБОЛИЗМ
Абсорбция
В силу своей химической природы свободный ион железа может быть в двух степенях окисления. Однако так исторически сложилось, что для организма не выгодно делать два аналогичных пути метаболизма, поэтому уже при поглощении железо поступает только в виде Fe2+. Как это происходит? Достаточно просто. Сначала все свободное железо просто окисляется до степени окисления 3+, а затем восстанавливается при поглощении до 2+. А теперь чуть подробней.
Первое соединение на пути свободного иона железа — редуктаза Dcytb (ферриредуктаза дуоденального цитохрома B). Перевод «ферриредуктаза» — восстановитель железа, напрямую говорит о функциях этого белка: он восстанавливает ион до формы Fe2+. В связи с этими функциями комплекс располагается на открытой в просвет кишечника части энтероцита, то есть на его апикальной мембране. Кроме ферриредуктазы восстановлением железа может заниматься и аскорбиновая кислота, которая попадает в кишечник в составе пищи. Об аскорбиновой кислоте и других стимуляторах абсорбции речь пойдет чуть позже.
Восстановление железа нужно для его прохождения внутрь клетки кишечника через белковый комплекс DMT1 (транспортер двухвалентных металлов).
Такая активная закачка железа происходит за счет параллельно происходящего транспорта протонов водорода, которые стремятся пройти через мембрану из-за накопленного на ней градиента, но сделать это они не могут в силу некоторых физиологических особенностей. Итак, DMT1 позволяет протонам делать то, что они хотят, а взамен использует энергию их перехода и вкачивает в энтероцит катионы железа. Что касается гемов, то они, попав в кишечник, тоже проникают в энтероцит*, но уже через другой трансмембранный белок —HCP-1 (белковый переносчик гема-1). В клетке гем распадается на железо и протопорфирин с помощью специального фермента HO-1 (гем оксигеназа-1). Восстанавливать железо до двухвалентной формы не нужно, так как оно уже находится в ней.
*Существует еще один путь поглощения гемового железа. Дело в том, что в селезенке содержатся специальные клетки-макрофаги, ответственные за перевариваривание старых и поврежденных эритроцитов. В макрофагах двухвалентное железо высвобождается из гемов гемоглобина и проходит тот же метаболический путь, что и в энтероцитах.
На этом этап абсорбции заканчивается. Ион железа полноценно поступил в организм и теперь у него есть три пути:
1.Пойти в расход на нужды энтероцита, то есть вступить во внутриклеточный синтез.
2.Если первый вариант не актуален и у клетки нет нужды в синтезе железосодержащих соединений, то запастись там же в составе белка ферритина, о котором мы еще поговорим.
3. Когда клеточных запасов будет достаточно, уйти в открытое плавание в кровоток для использования другими клетками организма.
Если ион железа решает пойти по третьему пути, то сначала он окисляется и транспортируется в кровь белком ферропортином и связывается там с апотрансферрином. Но об этом речь пойдет уже при обсуждении следующего этапа метаболизма-транспорта.
обобщенно стадия абсорбции железа выглядит следующим образом:
1. Железо высвобождается из соединений в пище с помощью протеаз и соляной кислоты. Соляная кислота желудочного сока может восстанавливать железо до Fe2+.
2. Свободные гемы попадают в клетку с помощью белкового переносчика гемов — 1 (HCP-1), расположенного в энтероцитах тощей кишки.
3. Внутри клетки гем диссоциирует на протопорфирин и двухвалентное железо благодаря гем оксигеназе.
4. Негемовое железо может реагировать с ингибиторами его всасывания в тонкой кишке, такими как полифенолы или щелочной pH, которые стимулируют фекальный вывод железа.
5. Редуктаза (например цитохром B) восстанавливает свободное трехвалентное железо до двухвалентной формы.
6. DMT1 переносит свободное двухвалентное железо внутрь энтероцита.
7. Fe2+ из свободного и гемового железа связывается с белками-переносчиками в цитозоле клетки кишки и либо используется клеткой, либо направляется в кровоток, либо на хранение в составе ферритина.
8. Ферропортин транспортирует двухвалентное железо сквозь базолатеральную мембрану энтероцита, после чего оно окисляется гефестином до трехвалентного состояния.
9. Две молекулы Fe3+ связываются с апотрансферрином с образованием трансферрина, который направляется к тканям организма через кровоток.
РЕГУЛЯЦИЯ АБСОРБЦИИ
Как вы, наверное, уже поняли, абсорбция железа- сложный многоступенчатый процесс, поэтому она может контролироваться на разных этапах и разными факторами.
Во-первых, активность абсорбции напрямую зависит от концентрации железа в межклеточном веществе вокруг энтероцитов, поэтому после приема пищи, богатой железом, ее интенсивность может резко упасть даже при дефиците микроэлемента в организме.
Во-вторых На поступление нового железа из кишечника влияют внутренние органы, которые могут стимулировать абсорбцию железа при недостатке микроэлемента с помощью гормона гепсидина. В частности, этим занимается печень: в ней железо запасается в виде составной части ферритина.
В-третьих, существуют отдельные уникальные регуляторные вещества, воздействующие на процесс поглощения либо гемового, либо негемового железа. Их делят на стимуляторы и ингибиторы в зависимости от своей функции..
СТИМУЛЯТОРЫ
Наиболее известный стимулятор абсорбции негемового железа — аскорбиновая кислота, или витамин C. Многие животные могут ее синтезировать, но приматам приходиться добывать ее с пищей. Как вы помните, аскорбиновая кислота восстанавливает трехвалентное железо, поэтому при ее избытке большее количество негемового железа способно поступить в энтероциты.
Но на этом эффективные стимуляторы абсорбции не кончаются:
Заменимая аминокислота цистеин. По сути тоже является органической кислотой. Входит в состав альфа- кератинов, участвует в синтезе коллагена и, соответственно, необходима для роста производных кожи: ногтей и волос.
Синтезируемый в теле человека трипептид глутатион, в состав которого входит и цистеин. Глутатион — это антиоксидант, вступающий в реакцию со свободными радикалами кислорода в клетке.
ИНГИБИТОРЫ
«Ингибитор» — полный антоним термина «стимулятор», ингибиторы снижают интенсивность абсорбции. В природе их существует огромное множество, но почему? Дело в том, что в основном это продукты вторичного метаболизма растений. Это значит, что растения не используют эти соединения для своего основного метаболизма и просто накапливают их, например для защиты от поедания. Вернемся к ингибиторам:
1. Полифенолы. Эти соединения содержится в больших количествах в продуктах растительного происхождения, таких как какао, чай, кофе, красное вино, зерновые культуры и тд. Наибольший эффект наблюдается при употреблении в пищу двух самых известных горячих напитков. Одна чашка кофе уменьшает эффективность усвоения негемового железа из мяса на 39%, а тот же объем чая — уже на 64%.
2. Фитаты. Производные фитиновой кислоты не усваиваются животными с простым однокамерным желудком, в том числе и человеком. Употребление этих соединений вызывает довольно сильное ингибирование абсорбции.
3. Кальций. Соли кальция также оказывают ингибирующий эффект, однако этот эффект уникален: в отличие от большинства ингибиторов кальций не специфичен к виду абсорбируемого железа. Это значит, что он в равной степени уменьшает всасывание как негемового, так и гемового железа. Эффект наблюдается только при одновременном наличии кальция и железа в просвете верхнего отдела тонкой кишки, а также при голодании человека.
Хоть рассмотренные стимуляторы и ингибиторы по своей сути являются физиологическими антагонистами, их силы все же не равны: наличие достаточного количества стимуляторов абсорбции пересиливает эффект даже больших концентраций его ингибиторов. Однако регулярное недоедание железосодержащих продуктов, стимуляторов абсорбции и/или обилие ингибиторов в пище может привести к дефициту железа. Отсутствие его запасов в организме, конечно, стимулирует абсорбцию, однако в данной ситуации не компенсирует эффект ингибирования полностью.
ТРАНСПОРТ
Вернемся к метаболизму железа. После поступления железа в энтероцит оно должно стать доступным каждой клетке тела, и в организме подобный транспорт осуществляется по кровотоку. Для реализации этой несложной махинации клетке кишечника нужно просто выбросить железо в кровь. На базолатеральной стороне энтероцита находится транспортер двухвалентного железа, называемый ферропортином, который и поставляет ионы в кровоток. Сразу после выхода из клетки мембраносвязанная ферроксидаза гефестин окисляет Fe2+ до Fe3+. Одной из возможных причин этого преобразования может быть разграничение пулов транспортируемого (3+) и используемого в клетке (2+) железа.
Затем железо попадает в плазму и следует до пункта назначения вместе со специальным кровяным транспортером — трансферрином (Tf). Белок связывает молекулы железа на двух сайтах и поддерживает их в нужной степени окисления. Доля железа, связанного с Tf, невелика — около 0,1%, но интенсивность его перемещения настолько высока, что каждый день подвергается более чем десятикратному обороту.
В свою очередь в процентном соотношении Tf тоже нечасто связывается с железом — только 10% от всех молекул. Такая форма трансферрина называется холотрансферрином (холоTf), остальной же несвязанный белок именуется апотрансферрином (апоTf). Такое высокое относительное содержание апоTf позволяет создавать кровеносный буфер для железа. Это значит, что ион, только что попавший в кровоток, сразу же связывается с трансферрином.
После связывания молекул железа из крови с апотрансферрином он превращается в холотрансферрин, который, в свою очередь, связавшись с мембранным рецептором к трансферрину (TFR), попадает в эндосому клетки, где высвобождает железо и переходит в свой несвязанный вид. После выхода трансферрина из клетки круг завершается и повторяется.
Что происходит дальше? Есть два пути: Большинство железа идет на эритропоэз в красном костном мозге, который использует исключительно Tf-железо. Другие органы и ткани также используют Tf-железо в своих
целях. Оставшаяся четверть железа запасается в организме, из которого почти 17% отправляется на хранение в печень. Второй путь мы рассмотрим чуть подробнее.
ХРАНЕНИЕ В ПЕЧЕНИ
Трансферрин путешествует по кровотоку в компании двух молекул трехвалентного железа. Его распознавание клетками печени, гепатоцитами, происходит с помощью мембранных белков — трансферриновых рецепторов (TfR). На месте связывания Тf образуется везикула, и белок-рецепторный комплекс попадает внутрь клетки. Протонный насос подкисляет ионами H+ среду эндосомы до pH 5,5, и молекулы железа отходят от трансферрина, который в свою очередь остается связанным с рецептором. Fe3+ восстанавливается до Fe2+ с помощью металлоредуктазы STEAP3 (Six Transmembrane Epithelial Antigen of the Prostate proteins 3) и в таком виде выходит из везикулы с помощью уже известного нам транспортера двухвалентных металлов DMT-1. Трансферрин же в своей апо-форме отсоединяется от TfR и экзоцитируется из гепатоцита в кровоток, а рецептор возвращается на мембрану клетки. Трехвалентное железо запасается в белковом комплексе — ферритине. При недостатке микроэлемента в крови гепатоциты начинают разрушать этот белок в лизосомах, высвобождают железо и секретируют его в кровь по схожему с энтероцитами механизму.
РЕГУЛЯЦИЯ МЕТАБОЛИЗМА ЖЕЛЕЗА
Метаболизм железа B организме контролируется в основном с помощью пептида гепсидина, секретируемого печенью. Ферропортин-единственный экспортер неорганического железа в теле млекопитающих. Гепсидин вызывает его инактивацию или даже уничтожение на макрофагах, энтероцитах, гепатоцитах и других клетках тела. Поэтому гепсидин является главным регулятором содержания железа в крови, с помощью которого организм изменяет доступность железа для тканей и регулирует скорость эритропоэза.
Заболевания с малым содержанием гемоглобина в крови, например анемия гипоксия, ингибируют синтез гепсидина еще на уровне мРНК в гепатоцитах. Результатом становится повышение общего уровня железа в организме, что в свою очередь стимулирует синтез гемоглобина.
Многие органы поддерживают постоянный уровень железа в крови через стимуляцию выброса железа печенью и его поглошения в тонкой кишке. Например, так делает красный костный мозг, так как ему необходимы ресурсы для самого затратного по железу процесса в организме - эритропоэза
ПАТОЛОГИИ
Ну и раз уж я заякнулась про всякие болячки, стоит сказать про них пару слов. Давайте для начала немного поясним терминологию:
симптом - это некий признак заболевания, а синдром - группа симптомов.
Анемии (малокровие) - группа синдромов, характеризующихся нарушением транспорта кислорода к тканям и развитием гипоксии.
Существует множество различных анемий (приведены далеко не все):
Железодефицитные анемии возникают из-за недостатка железа в организме. Такое может произойти как из-за малого содержания железа в пище, так и из-за проблем с его усвоением.
Дисгемопоэтические анемии возникают из-за нарушения образования эритроцитов в красном костном мозге.
Постгеморрагические анемии связаны с кровопотерей, например, после травмы или из-за паразита.
Гемолитические анемии возникают при разрушении эритроцитов, например, из-за змеиного яда.
Стандартной анемией, про которую вы наверняка уже знаете, является серповидноклеточная анемия.
Если определённый глутамат в В-глобине заменяется на валин, молекулы гемоглобина при потере кислорода агрегируют друг с другом в длинные фибриллы. Это приводит к изменению формы клеток: они вытягиваются, становятся менее гибкими и чаше застрявают в узких сосудах, что приводит к тромбозу.
Помимо серповидноклеточной анемии, с мутациями в гемоглобине связаны также талассемии. Гемоглобин состоит из двух видов цепей (а и В или а и σ).
Если поломать один из генов цепей, будут образовываться нестабильные полипептидные агрегаты из избыточных цепей, что в конечном итоге приведёт к лизису эритроцитов. Серповидноклеточная анемия - рецессивное заболевание. Гомозиготы по нему не выживают, а гетерозиготы в целом чувствуют себя нормально, пока не попадут в среду с недостатком кислорода (например, пока не полезут в горы).
Есть такой паразит из группы споровиков - малярийный плазмодий. Подробно обсуждать здесь его жизненный цикл не буду, однако кратко можно написать следующее:
Малярийный плазмодий проникает в эритроциты и размножается внутри них, тем самым пряча себя от иммунной системы. Когда приходит время, он перестривает актиновый цитоскелет эритроцита в своеобразные рельсы и пускает по ним свой белок - адгезин. Адгезин, выйдя на поверхность эритроцитов, заставляет их слипаться, а это приводит к лизису. Радостные малярийные плазмодики (и продукты их жизнедеятельности) выходят в кровоток, вызывая сильнейший иммунный ответ и лихорадку.
Но вот оказалось, что люди с серповидноклеточной анемией не
болеют малярией (даже если ей заражаются). Дело тут в том, что при наличии мутантного гемоглобина «рельс» из актина не хватает, чтобы довезти адгезин до поверхности, и малярийный плазмодий не может выйти из эритроцита.
Если поломать цитоскелет эритроцита (а именно, белок спектрин), то будет анемия Минковского-Шоффара. Эритроциты становятся шарообразными и теряют способность проходить через узкие сосуды.
Для синтеза эритроцитов необходимы различные вещества (в частности В12 и фолиевая кислота), и их дефицит также будет приводить к анемии.
Чтобы В12 (цианокобаламин) всосался в тонком кишечнике, он должен связаться в желудке с внутренним фактором Кастла. Если как-то поломать этот фактор, то развивается анемия
Аддисона-Бирмера. Лечение простое - колоть витамин В12 (как понимаете, пить его не выйдет).
РАСТЕНИЯ
На этом моменте мы радостно покончили с метаболизмом железа у животных, в частности у человека. Но я говорила, что железо также присутствует и в растительных тканях, ну как минимум в тех же Fe-S кластерах, между которыми перебрасывается этот несчастный электрон в световой фазе фотосинтеза.
В аэробных условиях и при рН почвы выше 6 Fe(II) в почве окисляется до Fe(III) и существует в основном в виде малорастворимых оксидов. В этих условиях концентрация растворимой формы железа обычно на порядки ниже концентрации, нужной растениям.
При дефиците железа у растения развивается межжилковый хлороз: промежутки между жилками желтеют, в то время как сами жилки остаются зелеными. Также замедляется рост и снижается урожайность.
В кислой среде концентрация растворимой формы железа повышается, как и его доступность для растений. Кроме того, большая его часть представлена Fe(ll) и может быть поглощена напрямую некоторыми растениями. Форма Fe(II) также более распространена в анаэробных условиях (например, на рисовых полях).
Для поглощения железа из почвы у растений есть 2 стратегии:
Первая характерна всем двудольным и однодольным, исключая злаки. Второй как раз пользуются злаковые.
В чем состоит первая стратегия:
Ну для начала неплохо было бы достать ионы железа из почвы, а учитывая, что оно там находится в основном в виде комплексов, сделать это достаточно трудно. Как действуют растения- они зачисляют pH среды в районе корня за счет H+/ATP-азы плазмалеммы, затем забирают железо из почвы, сразу восстанавливается, расположенной на мембране Fe3+хелатредуктазой (сопряжено с окислением NADFH), затем восстановленное Fe2+ переносится через мембрану специальным транспортером IRT1.
Растения со второй стратегией поглощают сразу Fe3+, они выделяют специальные низкомолекулярные соединения- фитосидерофоры (часто - небелковые аминокислоты, никотинамин), синтезирующиеся из метионина, содержащие три карбоксила, за счет которого они хелатируют железо.
Как понятно из определения фитосидерофоров, Fe3+ хелатируется внутрь такого соединения и вместе с ним переносится через мембрану специальным транспортером YSL.
Затем железо используется в растениях для производства различных соединений, например ферментов каталаз, пероксидах, таких соединений, как цитохромы, Fe-S кластеры и прочих других.
На этом увлекательный рассказ про железо подходит к концу, теперь вы знаете почти все об этом сложном микроэлементе. Надеюсь, вся эта информация о метаболизме железа, его регуляции в организме животных и растений была вам полезна, а если нет, то хотя бы интересна, приходите к нам в биологию :)