Это вторая часть конспекта видеолекции Павла Пашковского "ВЛИЯНИЕ КАЧЕСТВА СВЕТА НА РАСТЕНИЯ РОЛЬ ОСНОВНЫХ ФОТОРЕЦЕПТОРОВ И КОМПОНЕНТОВ ИХ СИГНАЛИНГА. PROlvl". Полный конспект с таймингом и сырыми данными ведётся здесь. Слова и изображения взяты из лекции. Источники - в конце.
Вопросы без ответов
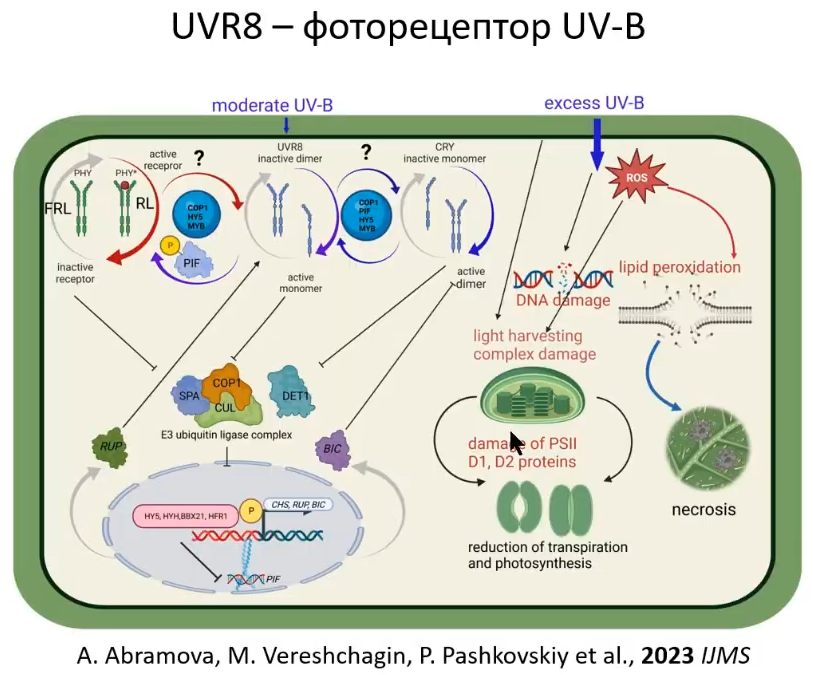
Этот слайд уже из нашей статьи. Он интересен тем, что по-прежнему даже в настоящее время при достижении больших результатов в этой области остаются вопросы. Например, один из интереснейших вопросов, которому мы обязательно посвятим часть своих работ: почему рецепторы, активируясь, превращаются в активные димеры?. Это связано и с криптохромами, и с фототропинами, и с фитохромами, но только не с рецепторами ультрафиолета B. Ультрафиолетовый рецептор активен в мономере.
Выглядят ультрафиолетовые рецепторы довольно забавно 😊: 2 тора соединены вместе, там образуются какие-то водородные связи, может, не напрямую, а через белки-посредники, а активация происходит именно разделением этого сдвоенного тора на 2. То есть, он разделятся при активации, а другие - соединяются.
Ещё один огромный вопрос: зачем нужны рецепторы ультрафиолета B? Я задавал этот вопрос и другим исследователям в области, в которой сам являюсь экспертом: ответа я получил пространных и 2. Первое предположение - это, что эволюция шла параллельно с эволюцией атмосферы, и в ранних слоях атмосферы проникало больше ультрафиолета и нужно было на него реагировать. Это объяснение было бы убедительно, если бы этот рецептор ультрафиолета B присутствовал во мхах, плауновидных, папоротниках, где этот рецептор отсутствует!
Точно сказать, есть он или нет, невозможно, необходимо проводить дальнейшие исследования. Я так понимаю, что есть какие-то папоротники, мхи, которые являются модельными организмами и их геном аннотирован, т. е. полностью прочтены последовательности их нуклеотидов, и среди этих последовательностей пока не обнаружена последовательность, кодирующая белки рецептора ультрафиолета.
Это первое хорошее объяснение, но оно не выдерживает критики.
А второе объяснение, которое может быть ещё лучше, но оно довольно пространное: не исключено, что рецепторы ультрафиолета - это молекула с более широким спектром действия, чем расчитываем, и свойства, которыми он обладает, ещё не открыты. То есть, мы знаем, что он реагирует на ультрафиолет B, а на что ещё он реагирует - мы пока не знаем.
Ещё есть такое предположение, что этот рецептор каким-то образом возник на будущее. Мы знаем, что растительные организмы очень пластичны и они способны приспосабливаться к абсолютно разым средам: могут расти и в горах, и при высокой температуре, и при высоком засолении. И, может быть, это такой запас свойств, которые растение закладывает для реализации в будущем.
Ещё один крайне интересный вопрос (в 2023 году я делал обзор литературы на эту тему - информации крайне мало): когда растение растёт под белым светом, активируются все фоторецепторы, а каким образом рецепторы взаимодействуют друг с другом?
Суть исследования - как взаимодействуют рецепторы вместе
Мы брали мутантов не по рецептору ультрафиолета, а по фитохрому и криптохрому (т. е. эти рецепторы были отключены), и смотрели, как ультрафиолет на них воздействует. Оказалось, что между рецепторами ультрафиолета и рецепторами красного света должна быть взаимосвязь при помощи факторов фитохромного сигналинга. Также, предположительно, что транскрипционные факторы MYB - это другие транскрипционные факторы, которые отвечают много за что, но, скорее всего, могут быть вовлечены. Взаимодействие уже между криптохромом и ультрафиолетовым рецептором, также, скорее всего, включает в себя все эти молекулы (COP1, PIF, HY5, MYB) и ещё что-то, что неизвестно (скорее всего, это компоненты светового сигналинга, о которых я уже говорил).
Так или иначе все виды спектра присутствуют в солнечном свете, и необходима координация между рецепторами, которые могут конкурировать за компоненты какого-то светового сигналинга. Т. к. они очень связаны, то получается, что некоторые белки, например HY5, присутствуют в световом сигналинге всех фоторецепторов. Поэтому, чтобы реакция растений была адекватной на световые условия, в которых растение произрастает, необходима взаимосвзь между фоторецепторами. Более того, фоторецепторы имеют очень большое взаимодействие с компонентами гормонального сигналинга, и здесь накладывается ещё более сложная регуляция, о которой можно отдельную лекцию рассказывать. Поэтому этот момент с взаимодействием фоторецепторов крайне не изучен.
F-box-белки
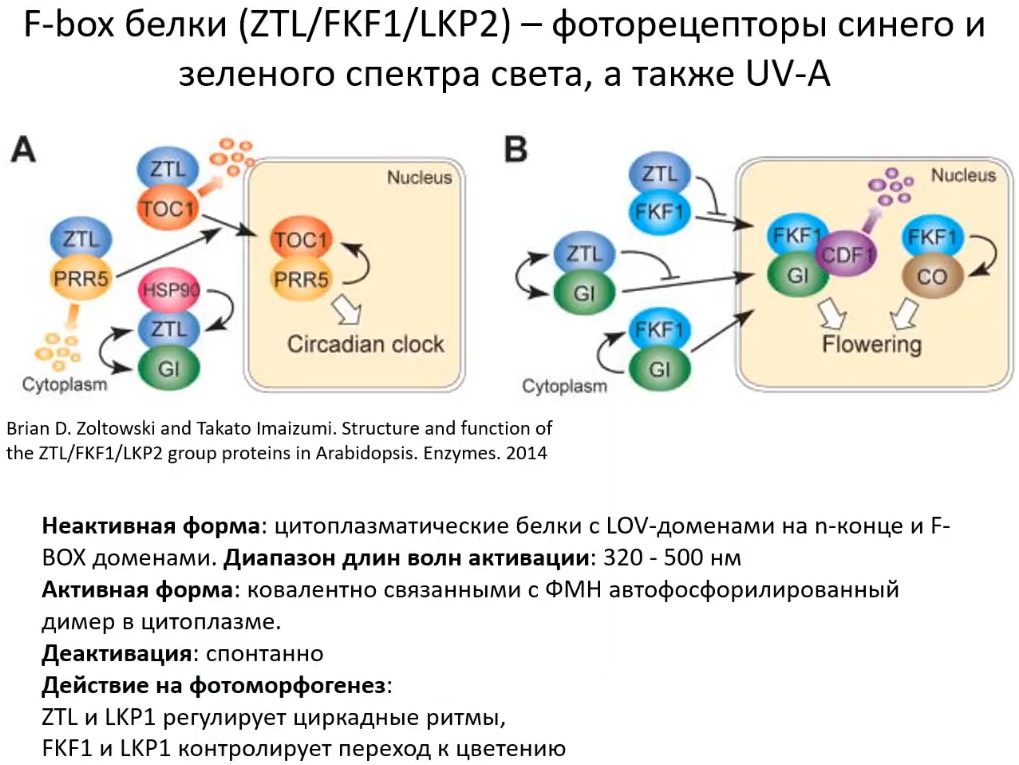
Я уже говорил про F-box белки (ZTL, FKF1, LKP2), так называемых фоторецептороподобных белках, у них также есть домены, чувствительные к свету и они способны также отправлять сигнал в ядро и участвовать в онтогенезе, переходе к цветению и многих других процессах. Фоторецептороподобные эти белки потому, что у них световая реакция проходит наподобие как в криптохромах: участвует ФАД, через ФАД происходит активация/деактивация этих молекул. Отличия - не все части рецептора, связывающиеся с ДНК, представлены как в классических рецепторах, т. е. они не могут весь ассамбль регуляций делать, способные на меньшее. Эти белки являются "рецепторами" не только синего, но и зелёного света.
"Рецепторы" активизируются при поглощении света и взаимодействуют с другими белками, трансфакторми, изменяют экспрессию светочувствительных генов. Несколько слов про то, что световые сигнальные пути имеют кросс-сигналинг между гормонами: криптохром, актвируясь, переходит в ядро и там способен взаимодействовать с промоторными областями ARF-белков. ARF-белки - это белки, которые участвуют в сигнале ауксинов. Также они могут быть регуляторами BZR-cигналинга, где могут в дополнение участвовать и белки PIF. Вместе с этим может быть регуляция гибберелловой кислоты (DELLA-белки). YUCCA - ауксиновый сигналинг, через ультрафиолетовый рецептор. TIR - непосредственно рецептор или часть рецепторов ауксинов, т. е. происходит ауксиновый сигналинг. При этом реакция происходит так, что фитохром реагирует не напрямую (как, например с ARF, который регулирует компоненты гормонального сигналинга), а через малые РНК (miR393) - это ещё один уровень регуляции, очень сложный, который направлен на включение определённых генов. Это когда из ДНК образуются РНК, а мы привыкли думать, что конечным результатом является белок. Так вот у некоторой части генов (у растения порядка 20% или 30% генов, у человека порядка 60%) конечной стадией работы является не белок, а регуляторная РНК, которая встраивается в специальный белковый комплекс, который несёт её к другой матричной РНК и способен либо временно остановить синтез белка, либо полностью разрушить матричную РНК и синтез белка вообще остановится. Матричная РНК кодирует белок (несёт информацию о последовательности аминокислот белка), а малая интерферирующая (микро) РНК выполняет регуляцию, и вот эта регуляция как раз и происходит через фитохромы. То есть на регуляцию, о которой я уже рассказывал до этого, плюс горманальная регуляция, накладывается ещё один уровень регуляции, выполняемый с помощью малых интерферирующих РНК. Я об этом тоже немного расскажу, потому что мы этим занимались в наших экспериментах.
Слайд говорит о светочувствительных процессах:
- фотоморфогенез: изменение формы и структуры растений,
- цикадные ритмы: внутренние биологические часы, синхронизация с физиологическими процессами продолжительности дня и ночи,
- фототропизмы:
1) движение опосредовано в направлении или от источника света,
2) избегание тени - это когда растения чувствуют соседей при помощи дальнего красного света. Когда дальнего красного много, то рядом много соседей, потому что все вкусные составляющие спектра (синий и красный), как правило, поглощаются хлорофиллами листьев выше растущих растений и в этот момент:
a) если это семечка, то семечка прорастает - задерживается на гетеротрофоном питании (питается за счёт эндосперма семядолей) и клетки не делятся, а растягиваются, повышается содержание ауксина, а также накапливается больше воды,
b) если это взрослое растение, то механизм примерно тот же, единственно, оно может замедлять (останавливать) фотосинтез и переходить к фотодыханию (неэффективный фотосинтез), потом об этом тоже скажу несколько слов, - синтез фитосинтетических пигментов
- синтез вторичных метаболитов
Есть фотосинтетически активная радиация, в которую вовлечены определённые спектры, а есть процессы, о которых я уже сказал до этого: например такие, как избегание тени - в них вовлечены другие части спектра, которые существенно меньше используются в фотосинтезе, а несут скорее сигнальную функцию, очень важную. Знание всего выше сказанного позволяет получать растения, более устойчивые и продуктивные, обладающие особыми, ценными для человека, биохимическими характеристиками, особенно в закрытых системах типа теплиц, сити-ферм и климатических камер.
Направления исследований
Объекты исследования
1. Arabidopsis thaliana - существует большое кол-во мутантов, которые можно использовать в работе. Это растение легко трансформируется, в него можно легко встроить любые гены. У него хорошо аннотирован геном (98%). Если исследовать тонкие моменты светового сигналинга, то использовать только Arabidopsis.
2. Томат - представлен: b - дикий тип, a - мутанты по основным фитохромам (A, B1, B2). У томатов фитохром B представлен двумя различными белками. Выполняет не только световой и температурный сигналинг, не только вовлечён в гормональный сигналинг и в антиоксиданты, но и ещё где-то, где растение решило сделать дополнительную копию белка. Различия не приципиальны, но есть. Это растение входит в десятку одних из самых аннотируемых геномов. Также это растение прекрасно клонируется, в отличие от Arabidopsis, что позволяет исключить изменчивость. Мутация заключается в том, что растение не чувствует свет и имеет специфические хлоропласты.
3. Сосны - важный хозяйственный объект: для строительства, получения различных материалов. У него есть целый комплекс недостатков: это перекрёстно- ветроопыляемое растение. То есть, найти одинаковые растения практически невозможно, клонированию подвергается очень плохо. Аннотация генома в плохом состоянии. Около 70-ти или больше хромосом, что очень большой геном. Слабые биохимические реакции (при существенном изменении освещения разница будет неособо заметна морфологически). Ещё у сосны есть своеобразные фитохромы: являются ортологами, то есть напоминают фитохромы высших растений, но одинаковыми не являются.
4. Маршанция - обладают рецептором фитохрома, который заключает в себе функции фитохромов обоих типов. Т. е. у него фитохром только одно типа.
Источники и персоналии
- Пашковский Павел, лектор конспектируемого видео, кандидат биологических наук, ведущий научный сотрудник лаборатории физиологических и молекулярных механизмов адаптации Института физиологии растений им. К. А. Тимирязева РАН. Специалист по фотоморфогенезу.