Мозазавр гоффмана (Mosasaurus hoffmanni) традиционно представляется как гигантский суперхищник, находящийся на вершине пищевой цепи. Однако, насколько это утверждение близко к истине? Сейчас, мы в этом разберемся.
1. Краткое описание
Mosasaurus hoffmanni - типичный представитель семейства мозазаврид (Mosasauridae): подсемейство мозазаврины (Mosasaurinae), отряд чешуйчатые (Squamata), крупнейший член своего семейства. Самым большим известным образцом может считаться образец из Пензы (Россия) - CCMGE 10/2469, с общей длиной черепа около 170 сантиметров (1700 миллиметров, как говориться в оригинальном исследовании от Grigoriev, D.V., 2014). Несмотря на изначально указанную длину в 17 метров, полученную по соотношению головы к телу 1:10, предложенному Расселом (Russell., 1967), более адекватные оценки образца говорят о максимальной длине в 13 метров, и массе около 5,5-10 тонн. Вид характеризуется небольшой глубиной морды и крупными зубами (Paul., 2022). M. hoffmanni существовал в период позднего мелового периода в маастрихтском ярусе около 70,5 миллионов лет назад (Polcyn et al., 2023) и имел космополитическое распространение по всей планете, распространившись за относительно короткий геологический период - около 25-30 миллионов лет (Lingham-Soliar., 1995). Судя по имеющимся данным, ящер достаточно успешно сосуществовал в одной экосистеме с другими крупными мозазаврами-макрофагами, не уступавшими, а иногда даже превосходившими его по размерам.
После такого небольшого экскурса можно приступить к полноценному разбору палеоэкологии мозазавра гоффмана.
2. Среда обитания
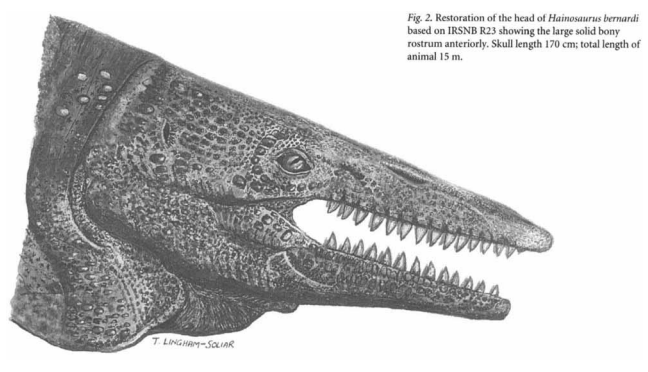
Судя по шельфовым залежам, моря маастрихтских отложений имели максимальную глубину в 300-400 метров, со средней глубиной около 50-и метров (Lingham-Soliar., 1995). M. hoffmanni обитал в прибрежных зонах, также предпочитая подобные места в виде охотничьих угодий, как показали анализы изотопов углерода δ13C. Схожую с мозазавром среду обитания предпочитали гализавр (Halisaurus) и гавиалимим (Gavialimimus), в то время как остальные районы (в том числе и мелководье) были заняты прогнатодоном (Prognathodon kianda; Strganac et al., 2015). Судя по всему, рядом с M. hoffmanni также сосуществовал крупный тилозаврин гайнозавр (Hainosaurus bernardi; Lingham-Soliar., 2007), который, вероятно, предпочитал схожие с мозазавром гоффмана среды обитания.
3. Пищевые предпочтения
Несмотря на гуляющее в рунете мнение о мозазавре, как об узко-специализированном макрохищнике, ряд анатомических особенностей не позволял M. hoffmanni быть эффективным охотником на крупную добычу. Несмотря на уменьшенный черепной кинетизм, мозазавр все ещё имеет весьма хлипкий челюстной сустав, что обуславливается квадратным мыщелком, создающим неглубокое сочленение с нижней челюстью. Современные крокодилы (Crocodilia), вырывающие куски мяса из тела жертвы, имеют куда более стабильное крепление челюстных суставов: их четырехугольный квадратный мыщелок идеально входит в углубление нижней челюсти. Это может говорить о том, что M. hoffmanni, в основном, проглатывал свою добычу, поскольку его крепление челюстных суставов позволяло ему очень широко раскрывать пасть для проглатывания относительно крупных кусков мяса. Избежать вывиха челюсти позволяли медиальные сухожилия, расположенные близко к суставу. В пользу проглатывания мозазавром добычи также говорит большой размер его головы. Несмотря на специфику строения челюстей ящера (в частности - относительно небольшой объем аддукторной камеры), M. hoffmanni был адаптирован к весьма высокой силе укуса (Lingham-Soliar., 1995). Сила укуса M. hoffmanni была рассчитана Benko., 2016 на основе промеров силы укуса аргентинских черно-белых тегу (Salvator merianae), имеющий схожее с мозазавром строение челюстных мышц и частичной реконструкции черепной мускулатуры M. hoffmanni от Lingham-Soliar., 1995. Как показали измерения, M. hoffmanni с 170-и сантиметровой нижней челюстью, кусал с силой в 110893 Ньютонов (11,3 тонн), что весьма впечатляюще для животного таких размеров.
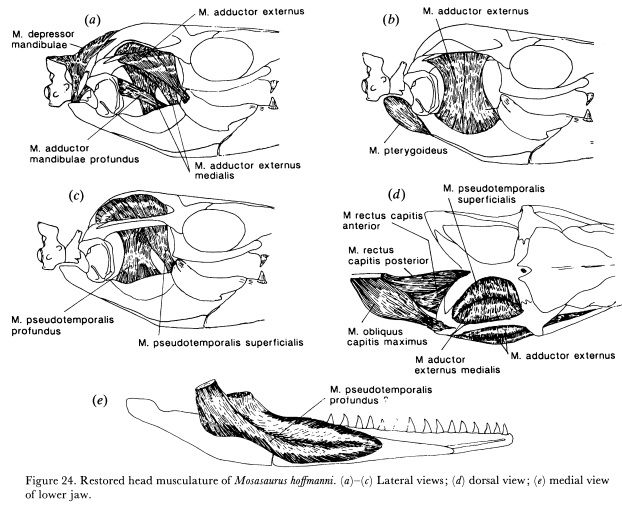
Свидетельств кормовой деятельности M. hoffmanni известно очень мало: нет ни копролитов, ни останков животных в области желудка. Имеются следы зубов мозазавра на 2,9 метровом панцире черепахи аллоплеврона (Allopleuron hoffmanni; ознакомиться с внешностью окаменелости можно здесь), но сейчас считается, что черепаха была атакована не M. hoffmanni, а крупным прогнатодоном (P. saturator; Dortangs et al., 2002). Однако, некоторые выводы можно сделать на основе близких родственников мозазавра гоффмана. Longrich et al., 2022, в своей работе с описанием талассотитана (Talassotitan atrox; о нём смотри ниже) упоминает, что Mosasaurus beaugei, морфологически очень схожий с M. hoffmanni (Street, H.P & Caldwell, M.W), был специалистом по питанию крупной рыбой. Кроме того, на специализацию к ихтиофагии у мозазавров подтверждается метровой рыбой в желудке 5,9 метрового Mosasaurus missouriensis (Konishi et al., 2014) и морфологией зубов различных мозазавров. К слову, именно зубная морфология даёт наилучшее представление о рационе мозазавра гоффмана: зубы M. hoffmanni чисто режущие, что может говорить о специализации на питании мягкой и костлявой добычей (Polcyn et al., 2006).
Все выше перечисленные особенности анатомии черепа M. hoffmanni, в сочетании с имеющимися доказательствами питания его близких родственников, позволяет сделать вывод о том, что жертвами мозазавра гоффмана являлись крупные рыбы (в том числе - небольшие акулы), в то время как на морских рептилиях хищничали в основном прогнатодоны (Prognathodon).
4. Взаимодействие с другими хищниками
Как уже упоминалось ранее, в одной формации с M. hoffmanni существовали несколько других крупных мозазавров: прогнатодон (P. saturator, P. sectorius), талассотитан (T. atrox) и гайнозавр (H. bernardi). Длительное выживание таких крупных хищников на одной территории может озадачить некоторых людей, однако здесь следует углубиться в экологию всех этих морских ящеров.
С P. saturator, P. sectorius и T. atrox все достаточно просто: они занимали ниши, отличные от таковой у M. hoffmanni, эксплуатировали иные среды обитания, как показали анализы изотопов углерода δ13C в зубах разных таксонов мозазавров (Strganac et al., 2015; Schulp, et al., 2013).
С H. bernardi ситуация несколько сложнее. Дело в том, что в работе от Lingham-Soliar., 2007 описываются черепные патологии молодого образца мозазавра гоффмана - IRSNB R12. В ходе исследования, автор приходит к выводу, что животное погибло преждевременно в следствие тупого удара чем-то узким по черепной коробке, как предполагается - рострумом гайнозавра (в статье проводится аналогия с современными афалинами (Tursiops truncatus), таранящими лимонных акул (Negaprion brevirostris)). Наличие подобного рода патологий у мозазавров гоффмана говорит о том, что даже такие крупные животные могли становиться жертвами других крупных хищников в молодом возрасте.
5. Палеопатология
Lingham-Soliar., 2004 описывает черепные патологии двух молодых образцов мозазавра гоффмана - IRSNB-25 и IRSNB-27. Образцы представлены частями нижних челюстей: челюсть первого (фрагмент зубной кости) была переломлена надвое в области нахождения шестого зуба. Стоит отметить, что на месте перелома начала образовываться большая костная "мозоль", на которой имелись следы бактериальной эрозии. Рядом с переломом также можно заметить несколько царапин. Второй образец также представляет из себя фрагмент зубной кости с некоторыми частями посткраниального скелета. Кончики передних зубов сломаны, и считается, что эти повреждения были получены животным при переломе. Кроме аналогичной предыдущему образцу мозоли, эта кость несет ещё два перелома, один из которых был почти полностью заживлен. Судя по всему, эти травмы не были смертельными, и животные прожили ещё некоторое время. Вероятно, что переломы могли сказаться на питании животных. Как показала аналогия с древесной агамой (Agama atricollis), серьезные нагрузки на челюсти при кормлении могли привести к деформации недавно сросшихся костей. Поэтому, мозазавры были вынуждены переместиться в более спокойную среду обитания, в которую не заплывают крупные животные (вроде взрослых особей M. hoffmanni или H. bernardi) и временно переключиться на питание более мелкими животными, благодаря своей способности глотать относительно большие куски мяса целиком. Причина переломов челюстей не ясна. Вполне вероятно, что описанные в работе образцы подверглись нападению более крупного образцы своего вида, что подтверждается аналогиями с современными рептилиями. Современные крокодилы имеют дело с агрессией со стороны сородичей (подробно эта тема разбирается тут; Somaweera et al., 2013), поэтому повреждения зубов для них - дело обычное. Возможно, для мозазавров были актуальны те же проблемы, что и для крокодилов.
6. Итоги
С учетом всего выше-перечисленного, можно сделать вывод, что мозазавр гоффмана не был высшим хищником даже для своего времени. Рядом с ним сосуществовали не менее крупные мозазавры, некоторые из которых могли охотиться на детенышей M. hoffmanni. Вероятно, что от представителей своего вида молодые мозазавры страдали не меньше, о чем говорят черепные патологии некоторых образцов. Краниальная анатомия, морфология зубного ряда и места обитания, с учетом имеющихся доказательств кормовой деятельности ближайших родственников, указывают на незамысловатую пищевую экологию одной из крупнейших хищных морских рептилий всех времён, с особым упором на питание животными небольших размеров.
Список литературы
1) Grigoriev, D.V., 2014. Giant Mosasaurus hoffmanni (Squamata, Mosasauridae) from the Late Cretaceous (Maastrichtian) of Penza, Russia. Proceedings of the Zoological Institute RAS 318(2):148-167
2) Paul, G.S., 2022. The Princeton field guide to Mesozoic sea reptiles. Princeton University Press.
3) Polcyn, M.J., Schulp, A.S., Goncalves, A.O., 2023. REMARKABLY WELL-PRESERVED IN-SITU GUT-CONTENT IN A SPECIMEN OF PROGNATHODON KIANDA (SQUAMATA: MOSASAURIDAE) REVEALS MULTISPECIES INTRAFAMILIAL PREDATION, CANNIBALISM, AND A NEW MOSASAURINE TAXON.
4) Lingham-Soliar, T., 1995. Anatomy and Functional Morphology of the Largest Marine Reptile Known, Mosasaurus hoffmanni (Mosasauridae, Reptilia) from the Upper Cretaceous, Upper Maastrichtian of the Netherlands. Philosophical Transactions of the Royal Society B: Biological Sciences, 347(1320), 155–172.
5) Strganac, C., Jacobs, L.L., Polcyn, M.J., Mateus, O., Myers, T.S., Salminen, J., May, S.R., Araujo, R., Ferguson, K.M., Goncalves, A.O., Morais, M.L., Schulp, A.S., de Silva Tavares, T., 2014. Geological setting and paleoecology of the Upper Cretaceous Bench 19 Marine Vertebrate Bonebed at Bentiaba, Angola. Geologie en Mijnbouw 94(01):121-136.
6) Lingham-Soliar, T., 2007. Unusual death of a Cretaceous giant. Lethaia, 31(4), 308–310.
7) Benko, D., 2016. Mosasaurus vs. Megalodon. Proza.
8) Dortangs, D.W., Schulp, A.S., Mulder, E., Jagt, J.W.M., Peeters, H.H.G., Th. de Graaf, D., 2002. A large mosasaur from the Upper Cretaceous of The Netherlands. Geologie en Mijnbouw 81(1).
9) Longrich, N.R., Jalil, N.-E., Khaldoune, F., Yazami, O.K., Suberbiola, X.P., Bardet, N., 2022. Thalassotitan atrox, a giant predatory mosasaurid (Squamata) from the upper Maastrichtian phosphates of Morocco. Cretaceous Research 140(10):105315.
10) Street, H.P., Caldwell, M.W., 2017. Rediagnosis and redescription of Mosasaurus hoffmannii (Squamata: Mosasauridae) and an assessment of species assigned to the genus Mosasaurus. Geological Magazine -1(3):1-37.
11) Konishi, T., Newbrey, M.G., Caldwel, M.W., 2014. A small, exquisitely preserved specimen of Mosasaurus missouriensis(Squamata, Mosasauridae) from the upper Campanian of the Bearpaw Formation, western Canada, and the first stomach contents for the genus. Journal of Vertebrate Paleontology, 34(4), 802–819.
12) Polcyn, M.J., Schulp, A.S., Mateus, O., Jacobs, L.L., Morais, M.L., Tavares, T.S., 2006. New mosasaur material from the Maastrichtian of Angola, with notes on the phylogeny, distribution and paleoecology of the genus Prognathodon.
13) Schulp, A.S., Vonhof, H., van der Lubbe, J.H.J.L., Janssen, R., van Baal, R.R., 2013. On diving and diet: resource partitioning in type-Maastrichtian mosasaurs. Geologie en Mijnbouw 92(2/3):165-170.
14) Lingham-Soliar, T., 2004. Palaeopathology and injury in the extinct mosasaurs (lepidosauromorpha, squamata) and implications for modern reptiles. Lethaia 37(3).
15) Somaweera, R., Brien, M., Shine, R., 2013. The Role of Predation in Shaping Crocodilian Natural History. Herpetological Monographs 27(27):23–51.