- Гипоксия, индуцируемый фактор и реакция на гипоксию.
Фактор, индуцируемый гипоксией (HIF), представляет собой фактор транскрипции, TOR , который служит главным регулятором транскрипции. реакции на снижение уровня кислорода
Специфичность специфичного для опухолевых клеток распознавания GAPDH HRE HIF-1 и HIF-2 была подтверждена в дальнейшем исследованиями на клеточных линиях рака молочной железы MCF-7 и SK-BR-3, предстательной железы и печени. Первоначальные исследования продемонстрировали индуцированную гипоксией экспрессию гена GAPDH. Увеличение мРНК и белка GAPDH, связанное с транскрипционной и трансляционной гипоксией, наблюдалось с помощью RT-PCR и иммуноблот-анализа соответственно. А6- и 2,5-кратное увеличение наблюдалось и в первом, и во втором. Кроме того, увеличение мРНК GAPDH зависело от времени, увеличиваясь последовательно через 12, 18 и 24 часа.
Kaelin, W. G. Jr. & Ratcliffe, P. J. Oxygen sensing by metazoans: the central role of the HIF hydroxylase pathway. Mol. Cell 30, 393–402 (2008).
Jiang, B. H., Rue, E., Wang, G. L., Roe, R. & Semenza, G. L. Dimerization, DNA binding, and transactivation properties of hypoxia- inducible factor 1. J. Biol. Chem. 271, 17771–17778 (1996). Seminal article on HIF.
- Гипоксия (дефицит кислорода) связана с увеличением генерации O2 ·- (и последующего H2 O2) вследствие ингибирования митохондриального ETC.
Hernansanz- Agustin, P. et al. Mitochondrial complex I deactivation is related to superoxide production in acute hypoxia. Redox Biol. 12, 1040–1051 (2017).
- В соответствии с этим было показано, что хроническая перемежающаяся гипоксия, опасное для жизни состояние, которое возникает при многих различных заболеваниях, включая апноэ во сне (нарушение дыхания во время сна), активирует окислительно-восстановительную передачу сигналов, что способствует нескольким системным и клеточным реакциям (которые включают изменения повышение артериального давления, повышенное высвобождение нейротрансмиттеров и нейротрофических факторов, а также изменение сна и когнитивного поведения), которые были связаны с активацией путей вторичных мессенджеров и регуляторов транскрипции гипоксии.
Prabhakar, N. R., Kumar, G. K., Nanduri, J. & Semenza, G. L. ROS signaling in systemic and cellular responses to chronic intermittent hypoxia. Antioxid. Redox Signal. 9, 1397–1403 (2007).
Оксиданты помогают стабилизировать HIF во время гипоксии, тем самым помогая вызвать гипоксическую реакцию.
Waypa, G. B., Smith, K. A. & Schumacker, P. T. O2 sensing, mitochondria and ROS signaling: The fog is lifting. Mol. Asp. Med. 47-48, 76–89 (2016).
Chandel, N. S. et al. Reactive oxygen species generated at mitochondrial complex III stabilize hypoxia- inducible factor-1alpha during hypoxia: a mechanism of O2 sensing. J. Biol. Chem. 275, 25130–25138 (2000).
- Пролилгидроксилазы HIF, которые определяют доступность кислорода и управляют гидроксилированием HIF и последующей протеасомной деградацией, модулируются оксидантами. Даже при нормоксии генерация оксидантов препятствует доступности Fe2+ в каталитическом сайте пролилгидроксилазы, ингибируя их активность и способствуя HIF-опосредованной транскрипции (см. reFs).
Pouyssegur, J. & Mechta- Grigoriou, F. Redox regulation of the hypoxia- inducible factor. Biol. Chem. 387, 1337–1346 (2006).
Acker, T., Fandrey, J. & Acker, H. The good, the bad and the ugly in oxygen- sensing: ROS, cytochromes and prolyl-hydroxylases. Cardiovasc. Res. 71, 195–207 (2006)
Таким образом, оксиданты влияют на пути HIF даже в негипоксических условиях, тем самым вызывая адаптивную реакцию на стресс.
- Регулирование датчиков стресса.
Тема гомеостатической адаптации, опосредованной окислительно-восстановительной передачей сигналов, также отражается в реакции различных сенсоров стресса на оксиданты.
Например, семейство транскрипционных факторов белка O (FOXO) способствует поддержанию клеточного и организменного гомеостаза посредством интеграции окислительно-восстановительных сигналов с другими сигнальными сигналами. Как наблюдалось в других окислительно-восстановительных сигнальных системах, это происходит посредством прямого окисления цистеина в членах FOXO, а также посредством окислительно-опосредованной настройки вышестоящих регуляторных механизмов.
Eijkelenboom, A. & Burgering, B. M. FOXOs: signalling integrators for homeostasis maintenance. Nat. Rev. Mol. Cell Biol. 14, 83–97 (2013).
Klotz, L. O. & Steinbrenner, H. Cellular adaptation to xenobiotics: interplay between xenosensors, reactive oxygen species and FOXO transcription factors. Redox Biol. 13, 646–654 (2017).
Существуют также прочные связи между оксидантами и р53, фактором транскрипции, который управляет ответами на различные стрессы, связанные с нестабильностью генома и нарушение регуляции которого тесно связано с раком. р53 находится под окислительным контролем, при этом H2 O2 модулирует селективную трансактивацию генов-мишеней р53, которая происходит опосредованно, через модуляцию сигнальных сетей, а возможно, и напрямую, за счет окисления остатков цистеина р53. И наоборот, р53 поддерживает клеточный окислительно-восстановительный баланс, регулируя экспрессию антиоксидантных генов.
Эта роль может поддерживать его функцию супрессора опухоли — в опухолевой нише, которая обычно является провоспалительной и характеризуется окислительным стрессом, р53 может ограничивать повреждение ДНК, вызванное окислительным стрессом, и прогрессирование опухоли. Регуляция энергетического стресса, опосредованная AMP-активированной протеинкиназой (AMPK), также находится под окислительно-восстановительным контролем.
Liu, B., Chen, Y. & St Clair, D. K. ROS and p53: a versatile partnership. Free Radic. Biol. Med. 44, 1529–1535 (2008).
Uehara, I. & Tanaka, N. Role of p53 in the regulation of the inflammatory tumor microenvironment and tumor suppression. Cancers (Basel). 10, 219 (2018).
Herzig, S. & Shaw, R. J. AMPK: guardian of metabolism and mitochondrial homeostasis. Nat. Rev. Mol. Cell Biol. 19, 121–135 (2018).
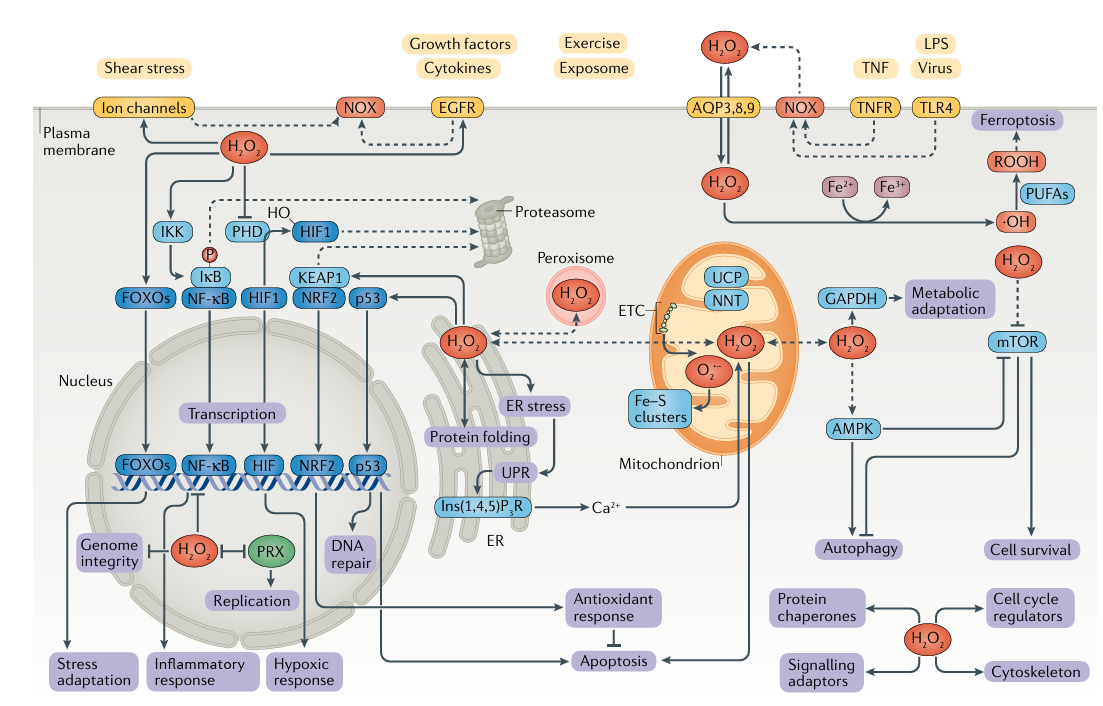
Однако, несмотря на то, что AMPK содержит остатки цистеина, которые могут подвергаться окислительно-восстановительной регуляции, недавно было показано, что активность AMPK в ответ на окислительно-восстановительные изменения не обусловлена прямым окислением самой AMPK, а является вторичным следствием окислительно-восстановительного воздействия на другие процессы, сходящиеся на других процессах. митохондриальное дыхание и приводит к снижению уровня выработки энергии.
Hinchy, E. C. et al. Mitochondria- derived ROS activate AMP- activated protein kinase (AMPK) indirectly. J. Biol. Chem. 293, 17208–17217 (2018).
Наконец, окислительно-восстановительная передача сигналов связана со стрессом, связанным с питательными веществами. Как избыток питательных веществ, так и недостаток питательных веществ связаны с повышенным образованием оксидантов и модуляцией функции ключевых сенсоров питательных веществ. К ним относятся AMPK (обсуждаемая выше), которая определяет доступность глюкозы, а также mTOR, которая реагирует на доступность аминокислот, стимулируя рост клеток, способствуя пролиферации и ингибируя аутофагию. Более того, аутофагия регулируется АФК как mTOR-зависимым, так и mTOR-независимым способом. Однако есть данные об канцерогенезе ,связанным с mTOR комплексом и ГАФДГ.
Liu, G. Y. & Sabatini, D. M. mTOR at the nexus of nutrition, growth, ageing and disease. Nat. Rev. Mol. Cell Biol. 10-0199 (2020).
Schmeisser, K. & Parker, J. A. Pleiotropic effects of mTOR and autophagy during development and aging. Front. Cell Dev. Biol. 7, 192 (2019).
- Глицеральдегидфосфатдегидрогеназа и метаболическая адаптация.
Последние исследования показывают, что GAPDH напрямую участвует в прогрессировании опухоли, ее инвазивности и метастазировании. Гипоксия индуцирует образование
Например, фермент ГАФДГ перемещается в ядро при окислении и выполняет функции репарации ДНК.
Другие негликолитические функции включают роль в экспорте тРНК и транспорте ER в аппарат Гольджи секреторного груза125. GAPDH также ингибируется H2O2, который реагирует с консервативным каталитическим цистеином, который необходим для ее чувствительности к гликолитическому субстрату, глицеральдегид-3-фосфату. Чувствительность этого цистеина к H2O2 по сравнению с глицеральдегид-3-фосфатом определяется ретрансляцией протонов с соседним цистеином, который, как было обнаружено, необходим для метаболической адаптации и выживания клеток в ответ на повышенные уровни H2 O2126.
- Эти исследования показали, что снижение GAPDH предотвращает апоптоз.
- Напротив, сверхэкспрессия GAPDH увеличивал скорость запрограммированной гибели клеток
- Эти исследования показывают, что GAPDH напрямую участвует в прогрессировании опухоли, ее инвазивности и метастазировании. В частности, эти исследования подчеркивают не только сложность ее канцерогенной активности, но и степень зависимости онкогенеза от этого ряда функций GAPDH. Таким образом, они указывают на значение и важность этого так называемого белка «домохозяйства» в отношении развития этой разрушительной патологии человека.
Разобщение белков и регуляция энергетики митохондрий.
Митохондриальная развязка — это процесс, при котором поток через ЦЭТ отделяется от синтеза АТФ. Это разобщение приводит к утечке протонов и, как следствие, к рассеиванию электрохимического протонного градиента в виде тепла, что является ключевым принципом термогенеза в термогенном жире (коричневые/бежевые адипоциты) и приписывается разобщающему белку, теперь называемому UCP1.
Lin, C. S. & Klingenberg, M. Isolation of the uncoupling protein from brown adipose tissue mitochondria. FEBS Lett. 113, 299–303 (1980). Discovery of uncoupling protein.
Echtay, K. S. et al. Uncoupling proteins: Martin Klingenberg’s contributions for 40 years. Arch. Biochem. Biophys. 657, 41–55 (2018)
Существует пять UCP, UCP1 USP5, и их роль выходит за рамки коричневой/бежевой жировой ткани, при этом разные изоформы экспрессируются в различных тканях.
Berry, B. J., Trewin, A. J., Amitrano, A. M., Kim, M. & Wojtovich, A. P. Use the protonmotive force: mitochondrial uncoupling and reactive oxygen species. J. Mol. Biol. 430, 3873–3891 (2018).
Jezek, P., Holendova, B., Garlid, K. D. & Jaburek, M. Mitochondrial uncoupling proteins: subtle regulators of cellular redox signaling. Antioxid. Redox Signal. 29, 667–714 (2018).
Разобщающей активности UCP способствует O2 ·-131, что вызывает легкое митохондриальное разобщение. Было показано, что это ограничивает активность ETC, тем самым обеспечивая отрицательную обратную связь на выработку митохондриальных оксидантов.
Кроме того, UCP2 и UCP3 поддерживаются в неактивном состоянии за счет глутатионилирования. Небольшое увеличение продукции митохондриальных оксидантов вызывает деглутатионилирование и тем самым активацию этих UCP. Кроме того, активность UCP2 включает окислительно-восстановительную активацию митохондриальной фосфолипазы, активность которой высвобождает свободные жирные кислоты, которые делают возможным UCP2-опосредованное разобщение.
Jezek, P., Holendova, B., Garlid, K. D. & Jaburek, M. Mitochondrial uncoupling proteins: subtle regulators of cellular redox signaling. Antioxid. Redox Signal. 29, 667–714 (2018).
Фосфорилирование и дефосфорилирование белков тирозина.
Протеинкиназы опосредуют различные клеточные процессы, начиная от пролиферации и дифференцировки и заканчивая принятием решений о выживании или гибели/апоптозе клеток путем фосфорилирования белков.
На фосфорилирование тирозина белка влияет прямая окислительно-восстановительная регуляция протеинтирозинфосфатаз и протеинтирозинкиназ.
Jezek, P., Holendova, B., Garlid, K. D. & Jaburek, M. Mitochondrial uncoupling proteins: subtle regulators of cellular redox signaling. Antioxid. Redox Signal. 29, 667–714 (2018).
Echtay, K. S. et al. Superoxide activates mitochondrial uncoupling proteins. Nature 415, 96–99 (2002).
Mailloux, R. J. & Harper, M. E. Uncoupling proteins and the control of mitochondrial reactive oxygen species production. Free Radic. Biol. Med. 51, 1106–1115 (2011).
С одной стороны, окисление реактивного цистеина в протеинтирозинфосфатазах вызывает их инактивацию, что приводит к повышению уровня фосфорилирования тирозина; Одним из примеров здесь является протеинтирозинфосфатаза 1B (PTP1B), окислительно-зависимая инактивация которой, как было показано, зависит от ее взаимодействия с белком 1433 .
Londhe, A. D. et al. Regulation of PTP1B activation through disruption of redox- complex formation. Nat. Chem Biol. 16, 122–125 (2020).
С другой стороны, протеинтирозинкиназы могут активироваться H2O2. Например, EGFR является мишенью сигнала EGF, полученного H2O2, а окисление цистеина активного центра до сульфената усиливает его киназную активность.
Truong, T. H. et al. Molecular basis for redox activation of epidermal growth factor receptor kinase. Cell Chem. Biol. 23, 837–848 (2016).
Аналогично, сульфенилирование цистеина приводит к активации киназы .
Heppner, D. E. et al. Direct cysteine sulfenylation drives activation of the Src kinase. Nat. Commun. 9, 4522–06790 (2018).
Было обнаружено, что CO2/бикарбонат необходим для окисления PTP1B, мишени в EGF-зависимой передаче сигналов H2 O2, демонстрируя роль HCO4– , а также CO2/бикарбонат усиливает опосредованное H2 O2 гиперокисление пероксиредоксина 1.
Dagnell, M. et al. Bicarbonate is essential for protein tyrosine phosphatase 1B (PTP1B) oxidation and cellular signaling through EGF- triggered phosphorylation cascades. J. Biol. Chem. 294, 12330–12338 (2019).
Truzzi, D. R. et al. The bicarbonate/carbon dioxide pair increases hydrogen peroxide- mediated hyperoxidation of human peroxiredoxin 1. J. Biol. Chem. 294, 14055–14067 (2019).
Помимо ферментов, адаптеры сигнализации также могут модулироваться посредством окислительно-восстановительной передачи сигналов. Например, GRB2-ассоциированный связывающий белок 1 (GAB1), многофункциональный адаптерный белок, играющий ключевую роль в сигнальных путях тирозинкиназы, недавно был идентифицирован как новая окислительно-восстановительная мишень NOX4 с Cys374 и Cys405 в качестве основных сайтов-мишеней.
Löwe, O. et al. BIAM switch assay coupled to mass spectrometry identifies novel redox targets of NADPH oxidase 4. Redox Biol. 21, 101125 (2019).
Кальциевые каналы ORAI, потенциалзависимые кальциевые каналы Нели и пуринергические рецепторы плазматической мембраны все они подвержены окислительно-восстановительной сигнализации.
Bogeski, I. & Niemeyer, B. A. Redox regulation of ion channels. Antioxid. Redox Signal. 21, 859–862 (2014).
Инозитол 1,4,5 трифосфатный рецептор (Ins(1,4,5)P3 R) Ca2+-каналы в ЭР также являются важными целями окислительно-восстановительной передачи сигналов и будет обсуждаться в следующем разделе.
Пероксисомальный H2O2 в окислительно-восстановительном гомеостазе и передаче сигналов.
Роль пероксисом в метаболизме липидов и H2O2 привела к новым интересным перспективам в передаче окислительно-восстановительных сигналов. Пероксисомы содержат ряд оксидаз, генерирующих H2O2 (таблица 1),
таких как жирноацил-КоА-оксидаза и оксидаза d-аминокислот, а также ферменты, восстанавливающие H2 O2, такие как каталаза и перидоксиредоксин . В то время как пероксисомальный метаболизм H2O2, очевидно, связан с регуляция пероксисомальных функций, он также воздействует на дополнительные пероксисомальные окислительно-восстановительные мишени, такие как FOXO3 или PTEN.
Fransen, M. & Lismont, C. Redox signaling from and to peroxisomes: progress, challenges, and prospects. Antioxid. Redox Signal. 30, 95–112 (2019).
Lismont, C., Revenco, I. & Fransen, M. Peroxisomal hydrogen peroxide metabolism and signaling in health and disease. Int. J. Mol. Sci. 20, ijms20153673 (2019).
Кроме того, пероксисомальные каталазы способны модулировать окислительный стресс на клеточном уровне, а каталаза даже может секретироваться, что связано со злокачественной трансформацией.
Gebicka, L. & Krych- Madej, J. The role of catalases in the prevention/promotion of oxidative stress. J. Inorg. Biochem. 197, 110699 (2019).
Böhm, B., Heinzelmann, S., Motz, M. & Bauer, G. Extracellular localization of catalase is associated with the transformed state of malignant cells. Biol. Chem. 396, 1339–1356 (2015).
Численность и распределение пероксисом сильно варьируют среди типов клеток, что повышает вероятность их важной роли в клеточно-специфической окислительно-восстановительной передаче сигналов.
Роль в сворачивании белков при стрессе ER и ER.
В контексте белкового метаболизма основным процессом является образование дисульфидных мостиков, которое для белков, поступающих секреторным путем, происходит в ЭР во время окислительного сворачивания белков. Для каждого дисульфида, образующегося в реакции, катализируемой протеиндисульфидизомеразами, образуется один окислительный эквивалент, H2O2, в результате повторного окисления протеиндисульфидизомераз, катализируемого эндоплазматическим оксидоредуктином 1 (ERO1). Следовательно, H2O2 является побочным продуктом сворачивания белка. Генерируемый ERO1 H2O2 может впоследствии использоваться глутатионпероксидазой , которая использует этот H2O2 для дальнейшего стимулирования окислительного сворачивания, в то же время снижая окислительную нагрузку на ER.
Wang, L., Zhang, L., Niu, Y., Sitia, R. & Wang, C. C. Glutathione peroxidase 7 utilizes hydrogen peroxide generated by Ero1alpha to promote oxidative protein folding. Antioxid. Redox Signal. 20, 545–556 (2014).
Различные типы клеток имеют разные секреторные продукты, зависящие от этих систем ЭР. Например, плазматические клетки используют H2O2 для поддержания выработки антител, а островки поджелудочной железы используют H2O2 для поддержания выработки инсулина.
Cenci, S. & Sitia, R. Managing and exploiting stress in the antibody factory. FEBS Lett. 581, 3652–3657 (2007).
Laporte, A., Lortz, S., Schaal, C., Lenzen, S. & Elsner, M. Hydrogen peroxide permeability of cellular membranes in insulin- producing cells. Biochim. Biophys. Acta Biomembr. 1862, 183096 (2019).
Дерегуляция окислительно-восстановительного баланса ER связана к активации развернутого белкового ответа (UPR), который в целом направлен на восстановление гомеостаза ЭР на стрессе. В частности, повышение уровня обоих окислители или восстановители в ЭР могут привести к Стресс ER и активация UPR. В свою очередь, активация Было показано, что UPR приводит к дальнейшему увеличению ROS. уровни, влияя на функцию митохондрий
Cao, S. S. & Kaufman, R. J. Endoplasmic reticulum stress and oxidative stress in cell fate decision and human disease. Antioxid. Redox Signal. 21, 396–413 (2014).
Eletto, D., Chevet, E., Argon, Y. & Appenzeller- Herzog, C. Redox controls UPR to control redox. J. Cell Sci. 127, 3649–3658 (2014).
В соответствии с этим взаимодействием между окислительным стрессом и ЭР стресс, эндотелиальная дисфункция, вызванная окислительным стрессом было показано, что ему противодействуют удары по UPR.
Amodio, G., Moltedo, O., Faraonio, R. & Remondelli, P. Targeting the endoplasmic reticulum unfolded protein response to counteract the oxidative stress- induced endothelial dysfunction. Oxid. Med. Cell Longev. 2018, 4946289 (2018).
H2 O2 и межорганеллярная сигнализация. Между передачей сигналов кальция и передачей сигналов H2 O2 происходят существенные перекрестные помехи175–177. Мобилизация Ca2+ из ЭР в ответ на стимуляцию факторами роста и гормонами, а также в ответ на стресс ЭР (см. ранее) происходит через каналы Ins(1,4,5)P3R, которые сами являются окислительно-восстановительными мишенями, и функционально соответствующие остатки цистеина в Ins(1,4,5) P3 R1 теперь идентифицированы.
Joseph, S. K. et al. Redox regulation of type- I inositol trisphosphate receptors in intact mammalian cells. J. Biol. Chem. 293, 17464–17476 (2018).
Примечательно, что ЭР устанавливает тесные связи (места контакта с мембраной) с другими органеллами, включая митохондрии, с функциональными последствиями; Места контакта ЭР с митохондриями играют важную роль в притоке Ca2+ в митохондрии. Было показано, что этот приток Ca2+ происходит в митохондриальных нанодоменах H2O2, которые индуцируются передачей сигналов Ca2+ на границе между ЭР и митохондриями.
Booth, D. M., Enyedi, B., Geiszt, M., Varnai, P. & Hajnoczky, G. Redox nanodomains are induced by and control calcium signaling at the ER- mitochondrial interface. Mol. Cell 63, 240–248 (2016). Description of H2 O2 redox nanodomains.
Csordas, G., Weaver, D. & Hajnoczky, G. Endoplasmic reticulum- mitochondrial contactology: structure and signaling functions. Trends Cell Biol. 28, 523–540 (2018).
Помимо митохондрий, ЭР также устанавливает места контакта с пероксисомами, и было высказано предположение, что эти три органеллы образуют «окислительно-восстановительный треугольник», который будет поддерживать обмен окислителей и регулировать окислительно-восстановительную активность в различных органеллах, тем самым действуя в качестве фокусной точки в окислительно-восстановительном процессе. обмен сообщениями.
Оксиданты в патофизиологии
Учитывая роль оксидантов в клеточных процессах, описанную в предыдущих разделах, неудивительно, что нарушение регуляции окислительно-восстановительного гомеостаза является распространенным патофизиологическим состоянием, обозначаемым как переход от окислительного эустресса к оксидативному дистрессу.
Egea, J. et al. European contribution to the study of ROS: a summary of the findings and prospects for the future from the COST action BM1203 (EU- ROS). Redox Biol. 13, 94–162 (2017).
Go, Y. M. & Jones, D. P. Redox theory of aging: implications for health and disease. Clin. Sci. 131, 1669–1688 (2017).
Здесь мы в качестве примера иллюстрируем передачу окислительно-восстановительных сигналов в норме и при заболеваниях .
Valko, M. et al. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 39, 44–84 (2007).
Milkovic, L., Cipak, G. A., Cindric, M., Mouthuy, P. A. & Zarkovic, N. Short overview of ROS as cell function regulators and their implications in therapy concepts. Cells 8, 793 (2019).
Выводы
В следующей статье представлю участие оксидантов, особенно H2O2, в некоторых органах с точки зрения их физиологии и нарушения регуляции болезненных процессов, подчеркивая их плейотропные, часто антагонистические эффекты.
Важно отметить, что на современном уровне знаний может быть сложно провести различие между полезной и вредной передачей сигналов (эустресс и дистресс).
Таким образом, потребуются крупномасштабные систематические исследования, чтобы определить взаимосвязи сигнал-реакция в каждом контексте, чтобы лучше понять сложную роль оксидантов в патофизиологии.