Всё началось в далеком 2005 г.. Тогда автор начал писать модель происхождения Жизни на Земле. Как уже в первые 50 млн лет, после охлаждения Земли, после конденсации воды, появилась удивительная молекула про РНК (про Рибонуклеиновая Кислота), из четырех компонентов (фосфорной кислоты, сахаров и пары нуклеиновых оснований) родился полимер - оказавшийся биополимером. Только этот полимер имел особое свойство - он имел свойства катализаторов и .. -он мог самовоспроизводиться. Правда не самопроизвольно, но был удобным инструментом-лекалом- материалом для этого. Но мы сейчас просто пройдем по следам истории -которая запечатлена в дискуссии на большом форуме, который был "утоплен" службами Запада, которые создавали информационный и интеллектуальный вакуум в бывшей СССР.
Дискуссию будем выдавать порциями, чтобы оценить ту информацию, которую тогда выдавал в общее пространство автор. И какие дискуссионные ответы он получал. Ну и примерим это к сегодняшнему-2025 г.
Псевдоним автора Астсергей |astsergey| (просто первые буквы Фамилии и имени)
Другие участники - Al1 - Александр
Итак , далекий 2006 г, когда автор уже имел написанную работу по Организации ДНК и происхождению Жизни:
стр1 / это страница на форуме в Мембране/
astsergey
Скрыть | 3 декабря, вс 2006 г., 23:27
Начнем с того , что теории генетики не существует. Есть структура ДНК . Есть модели клетки , ядра и органоидов - как гистологических объектов . Пептиды , как ни удивительно - наиболее старая , но все еще туманная страна , откуда постоянно появляются все новые и новые `инструменты` организмов и клетки . Только несколько месяцев назад луч света прорезал это `туманное царство`
http://bioinformatics.oxfordjournals.org/cgi/content/abstract/22/1/21
показав очертания некоих фундаментов их построения и функций . ДНК еще хранит свои тайны в простейшем коде A,T,C,G - где записаны технические библиотеки организмов . Библиотеки - этот термин наверное более подходит для описания существа функций ДНК . РНК, последние годы эти ранее неприметные - передатчики информации с ДНК кодов (месенджРНК) , тРНК ...и примерно все...- вдруг оказались гораздо более активными действующими лицами . Завтра продолжим.
--------------------------------------------------------------------------------
Al1
Скрыть | 4 декабря, пн 2006 г., 01:44
:) Ждём.
--------------------------------------------------------------------------------
astsergey
Скрыть | 4 декабря, пн 2006 г., 10:09
astsergey
Начнем с того , что теории генетики не существует.
=== РНК, последние годы эти ранее неприметные - передатчики информации с ДНК кодов (месенджРНК) , тРНК (транспортные), рРНК (рибосомные)...и примерно все...- вдруг оказались гораздо более активными действующими лицами . Праймеры или инициирующие векторы - `АДРЕСА` с помощью которых осуществляется адресное исполнение транскрипции ( биосинтез РНК на ДНК матрице) - были первые мРНК (микроРНК). Правда в клетках находили и другие мРНК ,но скорее считалось , что это `обрывки` от лизинга других более крупных РНК .
Липидные производные и другие классы более сложных биохимических агентов также играют существенную роль в организации клетки и ее органоидов .
Вся теория генома (генетики) практически состоит , подобно знанию алхимиков , из достаточно фундаментальных знаний об объектах исследования (выше-перечисленное) их структур , химических , биохимических параметров и в большей или меньшей степени их участия в жизни клетки и организма . Все-таки современная наука , другие области знаний : являются одним неиссякаемым источником информации для генетики . И всеми своими достижениями она обязана ,как никакая другая наука , другим областям знаний .
Открытие ДНК -
http://n-t.ru/tp/ns/bt02.htm
"...Началось все еще в прошлом веке, когда никому не известный швейцарский врач Ф. Мишер опубликовал в 1871 г. в берлинском «Журнале медицинской химии» свою знаменитую статью о выделении нуклеина из белых клеток крови больных. Слово это образовано от латинского «нукс» – ядро ореха, а окончание «-ин» подразумевало, что он содержит азот, то есть относился к азотистым веществам, подобно белкам.
В 1879 г. на нуклеин Мишера обратил внимание крупный немецкий химик К. Альбрехт Коссель, которого было бы вернее назвать биохимиком. Коссель выяснил причину подагры («боли в ногах» в дословном переводе), которая возникает в результате отложения в суставах нуклеина. Он открыл в нуклеине вещество желтого цвета, производное мочевой кислоты. Оказалось, что это гуанин, впервые выделенный в 1858 г. А. Штрекером из перуанского гуано – помета птиц, ценного азотного удобрения...."
--- ее двойной спирали :
http://4-03.olo.ru/news/comp/3255.html
"...25 апреля 1953 года в журнале Nature была опубликована небольшая статья Джеймса Уотсона (James Watson) и Фрэнсиса Крика (Francis Crick), в которой речь шла о структуре ДНК. Официально статьи такого размера считаются даже не полноценными научными публикациями, а лишь письмами в прессу. Однако именно с этой публикации письма началась, по большому счету, новейшая история генетики. В нем впервые зашла речь о двойной спирали ДНК...."
--------------------------------------------------------------------------------
astsergey
Скрыть | 4 декабря, пн 2006 г., 12:12
astsergey
astsergey
Начнем с того , что теории генетики не существует.
=== Все-таки современная наука , другие области знаний : являются одним неиссякаемым источником информации для генетики . И всеми своими достижениями она обязана ,как никакая другая наука , другим областям знаний .
Но вот удивительно , несмотря на все это , сами генетики - ревниво `охраняют` периметр своих интересов . А что охранять то ? Кроме уже гигантских библиотек томов с данными опытов - которым бы позавидовали и алхимики ( только их труды куда-то исчезли...)..??( Все-таки ,электронника , время - дают свои плоды - по крайней мере в части мультипликации бумаги...) И в которых - так и остались вопросы , которые должны бы были получить там ответ ?... Последние данные о `множественной` сущности информации в геноме - вообще лишают их надежды ,в обозримое время, прийти к чему-то обобщающему.... Эта `множественная` сущность и есть та огромная `сеть` связей - густой паутиной покрывающая геном и скрывающая истинные `кнопки управления` и процессинг в нем .
Даже точнее : не скрывающая , а открывающая как `матрешку` - множество сущностей и обратных связей , что ,конечно же , не упрощает задачу . До сих пор , основной мат.аппарат генетики (МАГ): статистика , и в последнее время задействована теория информации (ТИ). Результаты более чем скудные , т.к. статистика - не является методом продуцирующим теории ,даже более, иногда там скрывается сама теория (ввиду симметрии явлений ). ТИ хороша тогда , когда есть работоспособная модель . А сами генетики не совсем понимают как относиться к их объекту (геному)- как процессируемому или процессирующему ... или генерирующему сущности... Т.о., после трех десятков лет , уже активного исследования геномиальных аппаратов разных организмов и их компанентов , секвенсов геномов : нет математической модели организации процессинга в геноме . Ясно , что в такой ситуации , когда используя уже богатый опыт научного исследования объектов в разных науках , не удалось найти таковую , задача остается актуальной : надо искать адекватную модель .
Как ни странно , но иногда полезно вернуться к истокам . Сложность явления (процессинг в геноме), его явно многоуровневая система связей , многопластовые `мотивы` и явная предвходность ( зависимость от предыдущего состояния) - позволяют считать , что Реконструкция Эволюции Генома (РЭГ)- будет тем мат.аппаратом , который позволит заглянуть в `кухню` вышеописанных связей , понять движущие силы , а значит достичь их математического - адекватного описания .
Т.о. , может быть в первый раз(не считая ист.наук), наука возьмет свое начало от сущности явлений - т.к. там явно будут действовать фундаментальные законы Природы . Вот причина , по которой мы возвращаемся к началу Жизни на Земле ...Что бы понять , что ее двигало , усложняло и создало наблюдаемое нами ее многообразие ... А так же понять , что движет ею сейчас ....
--------------------------------------------------------------------------------
Al1
Скрыть | 4 декабря, пн 2006 г., 13:53
Пока всё просто интересно. Ждём продолжения. есть вопрос:
То, что программа - это не просто ДНК - это понятно, но вот что мне непонятно, считается, что всё закодировано именно звеньями ДНК, а последовательность звеньев не имет значения, не сильно мне верится в такое пренебрежение природой к сохранению информации, может именно последовательностью звеньев что то тоже копируется? скажем количество делений, или связи с мРНК.
--------------------------------------------------------------------------------
Al1
Скрыть | 4 декабря, пн 2006 г., 14:03
"звеньев что то тоже копируется?" =кодируется
--------------------------------------------------------------------------------
astsergey
Скрыть | 4 декабря, пн 2006 г., 14:25
Al1
что всё закодировано именно звеньями ДНК, а последовательность звеньев не имет значения,
--- наверное под `звеньями` имеете ввиду кластеры (гены) ?
не сильно мне верится в такое пренебрежение природой к сохранению информации,
--- Конечно последовательность важна , об этом впереди...
--------------------------------------------------------------------------------
astsergey
Скрыть | 4 декабря, пн 2006 г., 15:09
astsergey
Начнем с того , что теории генетики не существует.
=== Вот причина , по которой мы возвращаемся к началу Жизни на Земле ...Чтобы понять , что ее двигало , усложняло и создало наблюдаемое нами ее многообразие ... А так же понять , что движет ею сейчас ...В этом месте я ,обычно, привожу следующий пример :
Допустим , привезли современный компьютер в 1940 год дали его ученым ?.... они бы до сих пор `разгребали` этот продукт , в котором собралась целая эволюция решений: как хардверных , так и софтверных .... Даже с ошибками в хардвере , которые `нейтрализуются` софтвером (кстати о `мифе` совершенства геномов - организмов..)...
--------------------------------------------------------------------------------
astsergey
Скрыть | 4 декабря, пн 2006 г., 15:15
astsergey
..... Даже с ошибками в хардвере , которые `нейтрализуются` софтвером (кстати о `мифе` совершенства геномов - организмов..)...
---Кстати , тут есть безответный гол в ворота `Высшего разума`. Тот , кто создал жизнь ,если бы таковой существовал : конечно же не допустил бы такие `ляпы` или тут же бы их исправил - теми же средствами ...
--------------------------------------------------------------------------------
Al1
Скрыть | 4 декабря, пн 2006 г., 19:23
astsergey
astsergey
..... Даже с ошибками в хардвере , которые `нейтрализуются` софтвером (кстати о `мифе` совершенства геномов - организмов..)...
---Кстати , тут есть безответный гол в ворота `Высшего разума`. Тот , кто создал жизнь ,если бы таковой существовал : конечно же не допустил бы такие `ляпы` или тут же бы их исправил - теми же средствами ...
Ой не знаю. С теми знаниями о окружающем мире мы напоминаем рыбок, рассуждающих о том, есть ли жизнь вокруг аквариума. Давайте разбираться лучше с тем, что известно, и не лезть в тонкие миры.
К стати, ещё один вопрос, который не раз задавал многим генетикам, Но Вы единственный, кто положительно ответил на него, так - хотелось бы и продолжение услышать. Вопрос был такой: Существует ли какой либо механизм аварийного восстановления информации. но - в геноме.
--------------------------------------------------------------------------------
Al1
Скрыть | 4 декабря, пн 2006 г., 19:35
astsergey
Al1
что всё закодировано именно звеньями ДНК, а последовательность звеньев не имет значения,
--- наверное под `звеньями` имеете ввиду кластеры (гены) ?
Кластер это более новое название. Под кластером (любым) понимается нечто маленькое, однородное, объединённое в нечто большее.
не сильно мне верится в такое пренебрежение природой к сохранению информации,
--- Конечно последовательность важна , об этом впереди...
ок.
--------------------------------------------------------------------------------
astsergey
Скрыть | 5 декабря, вт 2006 г., 11:20
Al1
astsergey
.... Вопрос был такой: Существует ли какой либо механизм аварийного восстановления информации. но - в геноме.
---- Есть механизм `репаринга` - восстановления ошибок в ДНК . + сейчас открыта новая возможность : РНК -`восстановление` старой и уже не содержащейся в ДНК информации . Использованные мРНК (мессенджРНК) сохраняются в `р-телах` , при делении клеток части материнского клеточного материала делятся между двумя клетками и, видимо , они и являются необычными переносчиками `стертой`информации.
--------------------------------------------------------------------------------
Pashur Участник Клуба
WWW: 331505195
Скрыть | 5 декабря, вт 2006 г., 12:11
Al1
К стати, ещё один вопрос, который не раз задавал многим генетикам, Но Вы единственный, кто положительно ответил на него, так - хотелось бы и продолжение услышать. Вопрос был такой: Существует ли какой либо механизм аварийного восстановления информации. но - в геноме.
Репарация имеется в виду? Тогда в качестве именно "аварийного" варианта могу предложить SOS-репарацию. В более спокойных условиях - другие виды репарации.
--------------------------------------------------------------------------------
Al1
Скрыть | 5 декабря, вт 2006 г., 18:20
astsergey
...при делении клеток части материнского клеточного материала делятся между двумя клетками и, видимо , они и являются необычными переносчиками `стертой`информации.
Можно чуть поподробнее, или ссылку?
--------------------------------------------------------------------------------
Al1
Скрыть | 5 декабря, вт 2006 г., 18:23
Pashur
Репарация имеется в виду? Тогда в качестве именно "аварийного" варианта могу предложить SOS-репарацию. В более спокойных условиях - другие виды репарации.
1. где можно ознакомиться?
2. что значит "в более спокойных условиях"? как отличие от SOS реп.? или "не сдесь"?
--------------------------------------------------------------------------------
Pashur Участник Клуба
WWW: 331505195
Скрыть | 6 декабря, ср 2006 г., 14:16
Al1
Pashur
Репарация имеется в виду? Тогда в качестве именно "аварийного" варианта могу предложить SOS-репарацию. В более спокойных условиях - другие виды репарации.
1. где можно ознакомиться?
2. что значит "в более спокойных условиях"? как отличие от SOS реп.? или "не сдесь"?
Да, - "в отличие от SOS"
Введите в поисковик "SOS-ответ" - информации много.
--------------------------------------------------------------------------------
astsergey
Скрыть | 6 декабря, ср 2006 г., 22:02
Al1
astsergey
...при делении клеток части материнского клеточного материала делятся между двумя клетками и, видимо , они и являются необычными переносчиками `стертой`информации.
Можно чуть поподробнее, или ссылку?
--- Предлагаю пока верить , поищу (но мало времени ,постоянно в командировках). Информация точная , но механизм пока не ясен - поэтому я и предлагаю логичное и простое объяснение .
--------------------------------------------------------------------------------
astsergey
Скрыть | 6 декабря, ср 2006 г., 22:16
astsergey
astsergey
Начнем с того , что теории генетики не существует.
=== Вот причина , по которой мы возвращаемся к началу Жизни на Земле ...Чтобы понять , что ее двигало , усложняло и создало наблюдаемое нами ее многообразие ... А так же понять , что движет ею сейчас ...
---Позволю себе еще раз продублировать материал о происхождении Жизни на Земле (чтобы под рукой были все работы и была ясна нить логики , которой следовал автор при их написании ).
----Original Message----From: "Sergey Astashkin" <idsast000@mail.ru
To: ayotov@abv.bg
Date: Sat, 05 Jul 2003 19:18:22 +0400
Subject: origin of life-inglish
Зарождение жизни. 2002 г.
Используемые сокращения :
DNA- дезоксирибонуклеиновая кислота;
RNA- рибонуклеиновая кислота;
PUO(ПУО) - Пуриновые основания;
PMO(ПМО)- Пиримидиновые основания;
p-Nzd - про- нуклeозиды;
p-Ntd - про- нуклeотиды;
pNtd - полинуклeотиды;
Gl гликоли;
PAR про- реактор;
IE , ИЕ информационные единицы.
A-IB - блоки pNtd не содержащие информации .
Существующие модели появления жизни на Земле не отвечают на главные вопросы:
1-Движущие силы - `автопилоты` до появления клетки ?
2- Вероятность возникновения и путь?( Земля | да и Вселенная | слишком молода , чтобы обеспечить вероятность возникновения жизни из белков).
Предлагаемая модель ответила на эти вопросы. КАК же это случилось? Описываемые в работе химические реакции - конечно же являются одними из огромного числа , но выделяются мной , как наиболее вероятные и важные для описания процессов приведших к зарождению жизни на Земле. Основная идея , очевидность зарождения жизни - через информационную часть. Т.е. `цель` жизни и ее суть -самовоспроизведение , именно с него и началась история жизни на Земле. Очевидно , что DNA и RNA были основой возникновения клеточной структуры. Как и последующая эволюция: эволюция матрицы - DNA и RNA молекул. Как будет показано ниже , время возникновения первых про - DNA и про - RNA молекул , было исторически коротким. Вероятность возникновения жизни настолько высока , что придется принять, что Жизнь, в ее простейшем виде - самовоспроизведение по этой модели - может возникнуть всюду : где условия даже периодически повторяют `активные циклы` ( т.е. если даже основные температуры низкие , но существуют периоды с температурами : 10С < Т < 100С , с присутствием нижеперечисленных компонентов в атмосфере и на поверхности , и с ограниченными зонами перегрева ). В дальнейшем изложении своей теории, автор рассматривает только значимые для возникновения жизни процессы и реакции . В данном случае рассматриваются системы Земного типа.
----Можно классифицировать и эволюционировать историю развития Жизни на Земле :
А. Накопление простейшего органического материала ( углеводородов ).
Б. Накопление сложных орг.веществ в том числе гетеросоединений .Появление ПУО и ПМО и их концентрирование на силикатах и полифосфатах ;
С. Накопление p-Nzd и p-Ntd и их концентрирование. Адсорбция их на силикатах и полифосфатах ;
Д. Транскрипция поли-пронуклеотидов на них ;
Е. Увеличение степени полимеризации ,появление ИЕ;
--Примерный расчет:
Можно оценить поверхность про - реактора ( PRO-ALIVE Reactor- `PAR`) - который образовывался из уже известных нам пород , например : микропористого силикогеля ~ 10 кв.м./ ( гр.минерала ) или -10 degr(7) кв.м./(тон.минерала). Выбор минерала основан на близости структурных постоянных связей силиката к существующим комплементарным связям - 3,4 А / в действительности ,с учетом `экзотических `гликолей ,`работали` и другие размеры / :
Пример :
Силикаты Полифосфаты
Si-- O—Si Р-- O -- Р
I I I I
O O O O
I ~3,4A- I I~3,26A-I
РИС. 4
PUO- 3,4A -PMO (в полинуклеотидах)
I I
D.Ribos D.Ribos
\ /
HPO4
РИС.5
При величине природного минерального образования ( малого размера ) ~ до 100 тон
( около 7х6х7 метров) ( активной плотностью до 0,3), можно оценить поверхность одного PAR как -10 degr(9) кв.м.. При постоянной (-Si--O--Si-) , I ~ 3,4 А - получим плотность активных (РИС.3) центров ~ 3*10 degr (17)/кв.м.. Или для одного реактора, ~3*10degr(28) активных центров в виде ковров. Для заполнения одного такого - PAR необходимо ~ 3*10degr(28) молей нуклеиновых оснований (при заполнении 10% -3*10degr(27) молей , или около 2х тон смеси ( PMO и PUO ).
Такой реактор служит `фабрикой`, где идет самодоработка полинуклеотидов. Стерическое автопилотирование реакций полимеризации , так можно назвать эти процессы. Учитывая селективность процессов , необходимо признать , что предложенный механизм укладывается в требования логики и теории. Вероятность появления определенной последовательности DNA из 40 нуклеотидов ~ 4 degr(40) или ~ 10 degr(24) - переборов. Что может быть легко обеспечено уже при заполнении PAR. Можно принять что скорость обменов типа:
Si-O--PMO + PUO ---> Si-O--PUO + PMO
РИС. 6
Примем скорость обмена ~ 1 обмен/сек. Для нуклеида из 60 оснований - требуется вероятность ~ 4 degr(60) ~ 10 degr(36) , что обеспечится таким PAR за ~10 degr(9) сек или ~ 30 лет.
Ясно ,что остановки случайны, а обмены происходят циклически -`волнами`. Период волны ~ геологическим эффектам , например : волны от водоема (гейзеры и т.п.) + дожди. Получим min ~10 сек , max ~ 1час . Средний период ~ 30 мин. Т.о. в вышеописанном случае / скорость генерации для 60 оснований / ~ 30 лет х 1800 ~ 0,054 млн.лет .
Но учитывая поливариантность , видимо , этот срок ~ 100 раз меньше. Учитывая также низкий выход реакций / 3` - 5`/ ; и др. побочные процессы , конкурирующие реакции – уменьшение на 2 порядка , неселективность фосфатирования – уменьшение на 2 порядка , др. факторы 2 порядка / эта цифра увеличится ~ на 5 порядков , т.о. будем иметь ~ 50 млн.лет. Ясно, что такие системы будут `перегружены` бессмысленной информацией . Эти огромные довески , можно грубо оценить на одну IU ( про-ген ), как равному порядку поливариантности, т.е. приблизительно ~ 2-4 порядка . Для грубой оценки , можно взять величину гена нуклеазы 600 пар Ntd для него A-IB достигает от 60 000 пар до 6 млн.пар Ntd.
Ж. Появление `композитов`-шли естестевенно-случайные сборки липидов, нуклеотидов, полинуклеидов, металлокомплексов, аминокислот и других органических соединений в `композиты`- которые начинают участвовать в процессах обмена в этой `полинуклеидной каше.
З. Появление `свободных` полимерных молекул поли-продезоксинуклеотидов и поли-прорибонуклеидов накопивших необходимые ИЕ обеспечивающие цикл самовоспроизводства и "кооперация" с другими дополняющими ИЕ . Естественные оболочки - микропузырьки в минералах, были первыми `клетками`, в которых и обмен с окружающей средой иногда становился специфичным - по микротрещинам в ней. Пузырьки позволяли практически идеально `доваривать` смеси полинуклеотидов и органики попавшие в них. Можно представить, что такая микротрещина играла роль селективной мембраны. Т.к. ее размеры и состав селективно определяли скорость движения различных типов молекул. Не удивительно , что первые системы были `привязаны` к силикатам - еще и потому , что они легко и различными путями образуют пузырьки ( лава , вулканический пепел, перегрев силикатных растворов и т.п.) с большими удельными поверхностями, полузакрытого типа. Естественно ожидать, что первыми заработали силикато-активные органические `композиты` - они сразу изменяли ситуацию в `клетке` - позволяя изменять ее селективные свойства , селективно сорбировались на стенках `пузырьков.
С другой стороны, появляется другая схема эволюции полинуклеотидов.Эта схема не нуждается в построении ,тяжелой : практически и теоретически необъяснимой эволюции по Дарвину представляемую как линию непрерывного и постоянного увеличения `качества` материала. К сожалению , практически , трансформацию DNA невозможно включить в Дарвиновский механизм. Т.к. уровень мутационных возможностей в одном геноме сильно ограничен ,по крайней мере он не в состоянии произвести радикальную смену генотипа эволюционно (гладко) без общей радикальной смены геномиального инструментария. Второе, мы не наблюдаем таких линий в природе: ни в фауне, ни во флоре.
Автор считает, что описываемая выше схема - Твердофазной Матрицы -полностью соответствует требованиям параметров теории и параметров времени, необходимым для описания процессов, приведших к появлению живой материи на Земле. Эта модель дает пищу для понимания процессов эволюции и пути эволюции , в основе которой оказывается эволюция DNA и RNA. Самым неожиданным является вывод о гигантских `довесках` - бессмысленных ( не работающих ) последовательностей , что коренным образом меняет наши представления о пути будущей эволюции и самих механизмов эволюции .
Автор еще раз уточняет , что целью этой работы было: выяснить роль и возможность появления первичной `живой` структуры из автофункционирующего механизма, функционирующего на основе приемлемых начальных условий ( с химической точки зрения, геологической, физической и вероятностных критериев процессов , а так же логической достаточности и необходимости ).
Альтернативная эволюция реальней выглядит исходя из первоначальной `перегруженности` геномов , которые в процессе эволюции , при скрытых мутациях в зонах A-IB реализуют необходимый потенциал для трансформации DNA.Такой процесс является процессом увеличения `ценности` информации ,при уменьшении размеров DNA. Она не требует специального инструментария из гипотез , предполагающих практически осмысленное развитие. В ее основе простые принципы мутации перемещения , потери , замены, причем потери в размерах DNA неизбежны. Но повышение ценности информации компенсирует количественные потери, конечно же до известного предела.
----
--------------------------------------------------------------------------------
astsergey
Скрыть | 6 декабря, ср 2006 г., 23:20
astsergey
astsergey
astsergey
Начнем с того , что теории
генетики не существует.
===Вот причина, по которой мы возвращаемся к началу Жизни на Земле ... Чтобы понять , что ее двигало , усложняло и создало наблюдаемое нами ее многообразие ... А так же понять , что движет ею сейчас ...
--- После написания этой части и оценочных расчетов стало более определенной уверенность , что это и был путь Жизни . Но тогда ? Если на первом этапе была просто полимеризация (?)и еще не действовали биохимические ферменты и механизмы, то значит она и должна подчиняться законам действующих масс (?)
----
Исходя из вышесказанного и была написана следующая работа :
ЭВОЛЮЦИЯ
образование современных форм геномов
ВВЕДЕНИЕ
В этой работе проведен анализ последовательности процесса роста полимерных структур про-ДНК-РНК. В результате сделано заключение об организации ее - как первично-бинарной системы.
ОБЩАЯ ЧАСТЬ
На начальной стадии развития живой материи на Земле происходил рост полимерных структур из смеси мономеров . Естественно принять , что синтез последовательностей из нуклеиновых оснований на этой стадии описывался обычным кинетическим уравнением . Закон
этого роста можно описать с помощью гипотетического уравнения (1) полимеризации :
(1) a*A + b*B + c*C = AaBbCc ...
Тогда скорость полимеризации представится следующим образом :
(2) V = K*[A]^a * ^b * [C]^c ...
Где : V – скорость роста полимерной цепи из мономеров видов - A, B, C.. ;
K – коэффициент ;
[A], , [C]...- соответствующие концентрации ;
a, b, c... - стехиометрические коэффициенты.
Ясно , что нас интересует разница в скоростях роста в естественных условиях , когда концентрации оснований нестихеометричны. Для того чтобы исследовать поведение реакции полимеризации в этих случаях возможно принять следующие допущения :
[A] = = [C] .. = [S] и
a = b = c .. = i/N
тогда :
(3) V(N) = K*[S]^i ;
Где :
[S] = гипотетическая средняя концентрация мономеров ;
N = число видов нуклеиновых оснований ;
i = длинна про-ДНК-РНК полимера
Когда одна из концентраций будет существенно меньше других ( например , примем – 0.5 * S ) , тогда получим :
для : N(2)=2 и N(4)=4 , когда [S] = 1 при длинне ДНК-РНК = 100 оснований .
Если одна концентрация будет : [C]=[0,5*S] , получим :
R2/4= V(2)/V(4)= K*[S]^100 / ( [S]^75 * [0,5*S]^25 } = 1 /(0,5^25)
Где :
R2/4- отношение скоростей полимеризации : для N(2)=2 и N(4)=4 .
Тогда :
R2/4 = 1 / (0,5^25) ~ 3,3 * 10^7
Если данная концентрация будет меньше других ( например 50% [0.5*S] от других ) отношение скоростей полимеризации при прочих равных условиях будет ~ 3.3 * 10^7 как было бы , например , при вариантах : `A`+`T` и `A`+`T` + `C`+`G` или на семь порядков меньше .Таким образом,практичeски не были бы воспроизведены пары , как в нашем случае `C` +`G`. Принимая во внимание трудности связанные с естественным синтезом и стабильностью`C` , сделанное предположение является правдоподобным , в предположении
естественного происхождения `C` . Таким образом ясно , что скорость полимеризации в случае двойничной модели будет на 7 порядков больше , так что, необходимо принять , первичные (архаичные) полимерные проДНК-РНК структуры - как бинарные комплементарные системы . Здесь возникает одно `НО` - мера архаичности ? Автор не считает , что простейшие (микробы) обязательно архаичны , более того , большинство из них новейшие простейшие .
Статистический анализ было бы логично делать не на основе отношений `A`+`T`/`C`+`G` , а по длине мултиплетов (A..),(T..),(C..),(G..)(A/T..),(C/G..) - например :АААААА. - А(6) – мультиплет .
Так что , там где статистический смысл последовательностей начинает исчезать и возможно с высокой вероятностью считать их рудиментами. С другой стороны , в результате полиморфизма изофункциональных кластеров ДНК , структура также несет в себе ее историю происхождения и глубину эволюции кластеров , геномов . Ясно , что эволюционно , последующее появление еще одной пары : ` C`-`G` , позволило увеличить информационную емкость проДНК-РНК при той же термодинамической устойчивой длине . С точки зрения последующей трансформации РНК , необходимо признать , что эволюция нашла еще один способ `уплотнить ` ДНК информацию. Поэтому ,следует ожидать что в ( в отдельных геномах до 2-4 ) геномах , будет `чувствоваться` старая ДНК ( как `платформы` ) на которой шли активные мутационные процессы . Но это стало возможным с появлением ` собственного ` производства `- ` C` и `G `.
--------------------------------------------------------------------------------
astsergey
Скрыть | 6 декабря, ср 2006 г., 23:30
----Продолжение
СРАВНЕНИЕ МУЛЬТИПЛЕТОВ ГЕНОМОВ .
Таким образом, предлагается модель развития первичных живых матриц (PAM – Primary Alive Matrix ) где : `AT `-матрица была основной и древнейшей. Сохранены и биохимические ` START ` - `AUU ` и `STOP`-`UAA` кодоны . Матрица `CG` - появляется позже , когда появляется биохимический синтез этих нуклеиновых оснований . Очевидно , что когда синтез появился :`C` и `G` – начали замещать `A` и `T`- генетический материал. `CG` - матрица входила в древний геном сначала как несущественная ошибка .И наиболее древние копировали ее чисто механически. Но , когда источник – биохимические циклы синтеза `C` и `G`появились , их присутствие уже не лимитировало ( как другие ошибки ) развитие и самовоспроизводство. И новая пара нуклеиновых оснований нашла свое место в эволюции . Ясно , что мы находимся далеко от этой удивительной эпохи – ДВОЙНИЧНОЙ ЖИЗНИ . С другой стороны , мы можем наблюдать сложные эволюционные процессы :
1. Распад `AT ` платформ ,
2. Замещение `AT ` платформ - `CG ` платформами .
3. Вырождение `AT ` - платформ .
4. Вырождение `CG ` - платформ .
Распад матриц мы можем определить из следующих соображений :
Примем : Скорость распада мультиплета размера (N) - зависит от его связанности с геномом :
(4) dN/dt = F(N)
Где: N – размер мультиплета .
F(N) - функция зависящая от `N` размера мультиплета и его `связанности`с геномом .
t - `нормализованное`время.
Скорость распада количества мультиплетов - (n) пропорциональна их количеству :
(5) dn(N) / dt = Kn * n(N)
Где: n(N) – количество мультиплетов с `N` – размерами
Kn – коэффициент .
Используя уравнения (5) and (6) получаем :
(6) dN = F(N) * dn/(( Kn )* n(N) )
Функция F(N) – зависит от степени участия мультиплетов в жизненно-важных
кластерах : как `ключей ` , ` инициирующих последовательностей` и знаков препинания в `грамматике` генома . Можно ожидать три простейших случая :
(7) а) F(N) = KN* N - когда мультиплеты размера -`N` не связаны своим
размером с функциями генома ;
Где: KN – Коэффициент характеризующий связь мультиплетов `N` – размера с геномом ( связь слабая ) ;
б) F(N) = KN – когда мультиплеты `N` – размера связаны своими кодирующими свойствами с геномом ;
в) F(N) = KN / N - когда мультиплеты размера `N` сильно связаны своим размером с функциями генома и их приемлимая для генома мутация пропорциональна их размеру ;
Соответственно получаем основные зависимости `N` от` n(N)` :
(8) для 7 а Ln ( Ni/Nk ) = K1 * Ln ( n(Ni) / n(Nk) )
(9) для 7 б Ni -Nk = K1 * Ln (n(Ni) / n(Nk) )
(10) для 7 в Ni2 -Nk2 = K1 * Ln (n(Ni) / n(Nk) )
Где : К1 = KN / Kn
Можно ожидать , что разные размеры и виды мультиплетов будут находиться в
различной зависимости от генома и ,соответственно , описываться различными уравнениями . Эти зависимости позволяют измерять `дискретное` значение скорости мутаций в отдельных участках кластеров геномов . Ясно , что эти значения носят вероятностный характер и требуют дополнительных подтверждений. Обычно функция F(N) - имеет отрицательное значение , при ее положительном значении мы будем иметь рост размеров мультиплетов .
СТАТИСТИКА
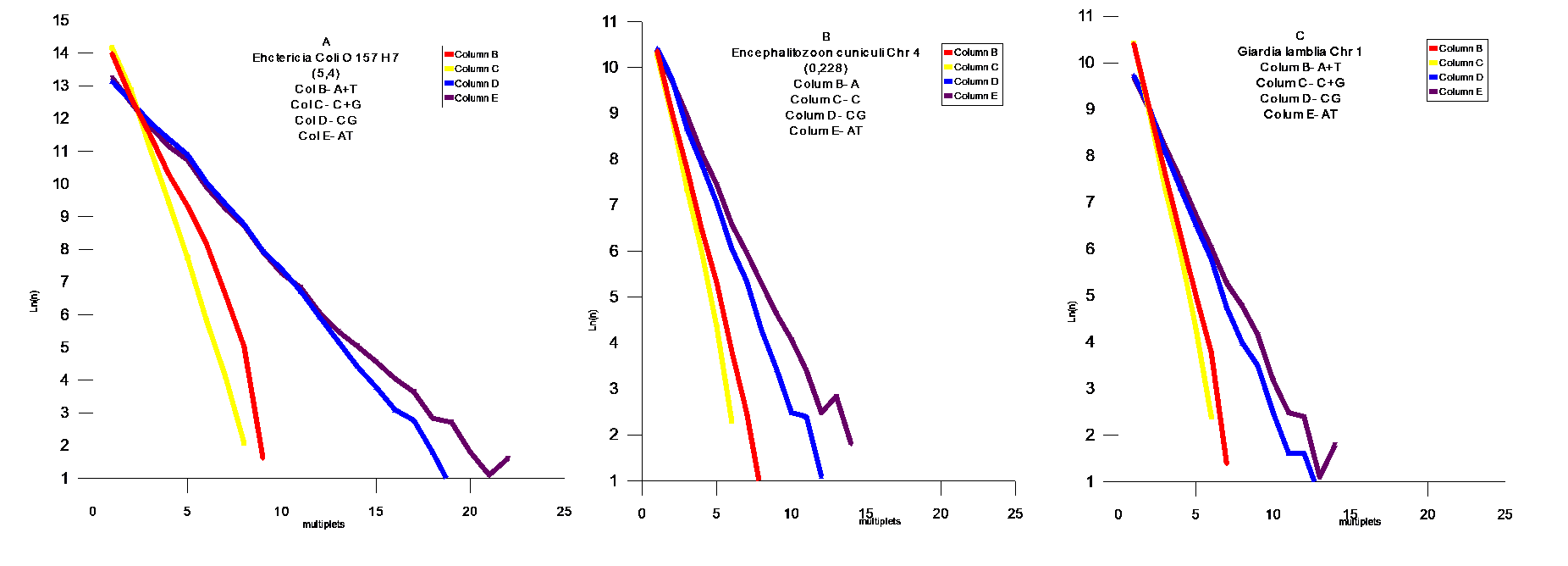
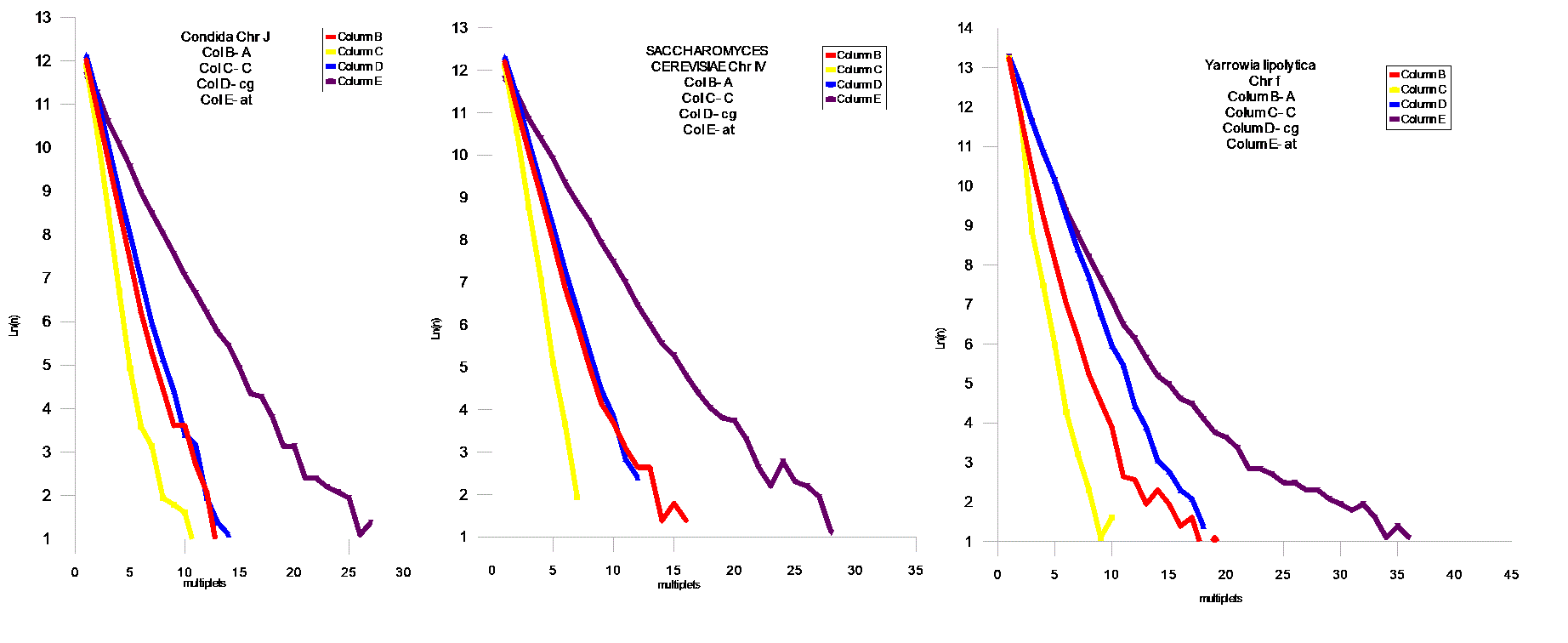
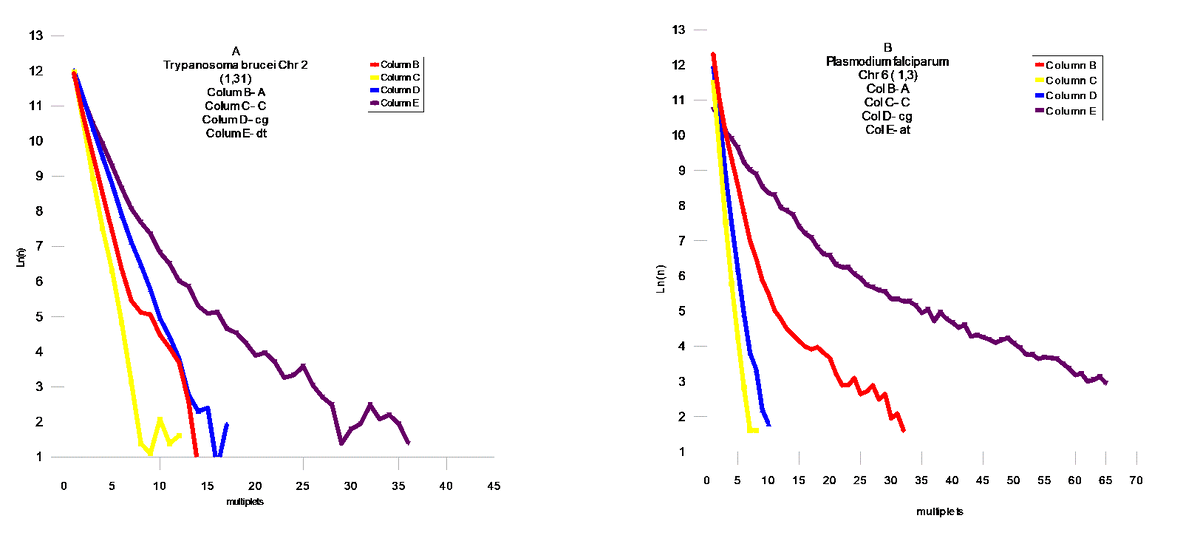
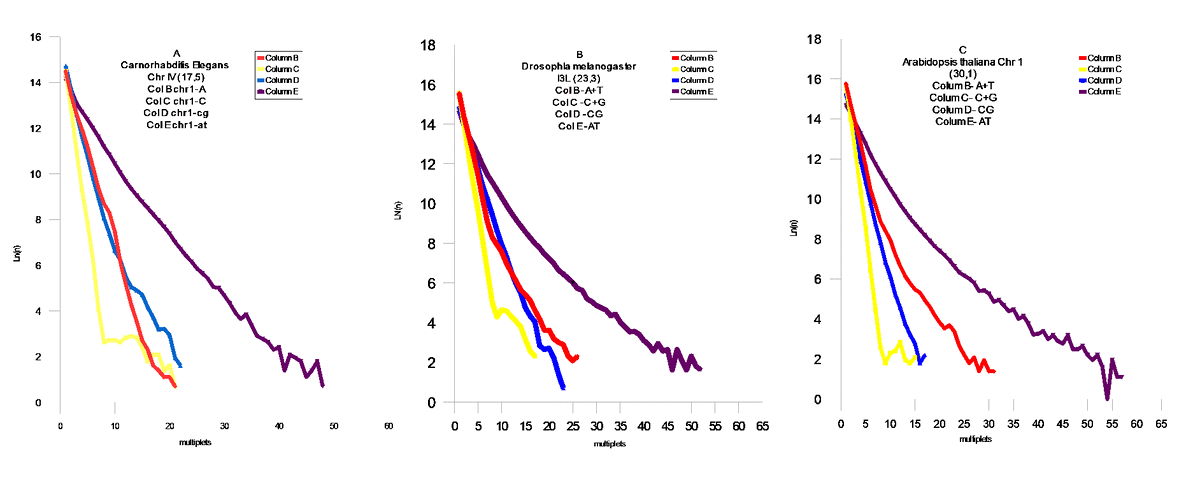
Проведен статистический анализ геномов из различных классов живых организмов на присутствие мультиплетов из `A` ,`T` ,`A`+`T`,`C`+`G`,`AT`, `CG`. Статистический анализ проводился подсчетом мультиплетов ( например : `А` – сумм ААА , `A`+`T` and `C`+`G` - простой суммой соответствующих размеров мультиплетов `A` и `T`, `C` и `G`- соответственно . `AT` и `CG` - заменой `T`на `A`, и `G` на `C` соответственно в исследуемом геноме .
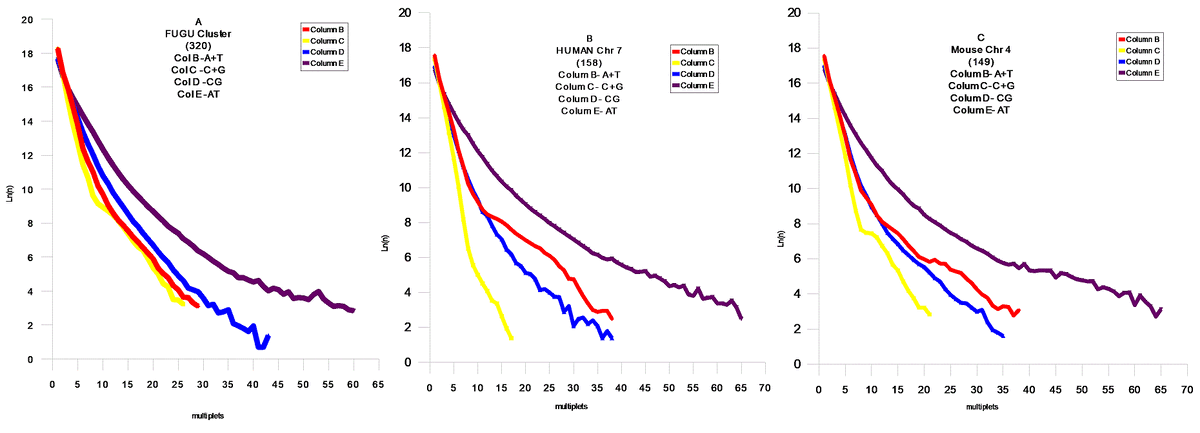
FIG.4 Middle Modern I , A- Caenorhabditis Elegans Chr IV,B- Drosophila melanogaster Chr 3L , C- Arabidopsis thaliana Chr 1 , FIG 5 – Fugu cluster .
FIG.5 Middle Modern I , B- Homo Sapiens Chr 7 , C- Mouse Chr 4 .
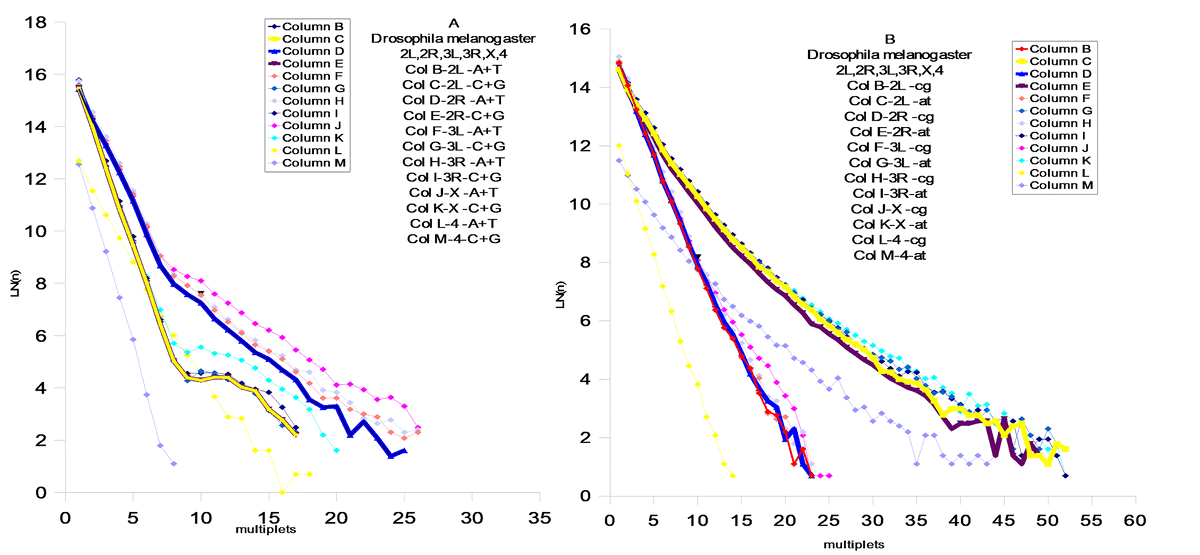
FIG.6 Drosophila melanogaster – Chr 2L, 2R , 3L , 3L , X , 4
FIG.7 – Examples of approximation for decayed vectors .
http://f.foto.radikal.ru/0610/2fec310efee5.jpg
http://k.foto.radikal.ru/0612/518b699d03c9.jpg
http://k.foto.radikal.ru/0612/c87bff34eb12.jpg
Модель поведения AT и CG матриц
В соответствии с результатами статистической проверки предложенной `AT`- модели , были обнаружены `AT`- матрици. Наблюдается тенденция `сжатия`пространства между `AT` и `CG` - матрицами . Соответственно , мы можем наблюдать этот процесс в развитии для различных геномов . Отмеченный процесс развития геномов дает векторы их развития, которые позволяют делать вывод об относительном эволюционном уровне генома . Модель позволяет сделать генетическое сравнение развития организмов из различных классов Царств Животных и Растений. Становится возможным установить их иерархию .Скорости распада и замещений отдельных групп дают возможность расчитать региональные скорости мутаций и их векторы .Хромосомные геномы показывают коллениарное поведение хромосом . Мы наблюдаем `дирижируемое` поведение мутаций в хромосомах одного генома. На Фиг.8 показана хромосома Leishmania major (а) и Leishmania infanta (б) . Это представители новейшего типа генома , который явно образовался путем `варки` в бульоне из разрушенных ядер из представителей современных ( modern) геномов - `вивисекцией` существенных кластеров , и то `АТ ` - содержащих : например с помощью` тепловой ` обработки . Причем Leishmania infanta явно показывает тотже принцип сжатия AT – CG пространства ,но уже с `другой` стороны .
Выводы
Эволюция геномов явно указывает на общий довлеющий фактор – увеличение компактности информации в геноме . Это достигается в основном уравновешением соотношения A,T / C,G . Сам механизм `мягкого` воздействия этого фактора , лежит явно в кинетике процессов воспроизводства . Степень `информационного` совершенства : это приближение этого отношения к `1` . Кроме того , размер мультиплетов стремится к равновесным ( вероятностным) значениям мультиплета (A,T,C,G).То есть , всякий геном имеет идеальный размер основания `m`- в зависимости от его размера `N` . Соответственно можно предложить меры идеальности геномов . Первая должна отвечать за `Первый` уровень совершенства когда отношение C,G/A,T – стремится к `1` , для этого удобно сравнивать суммы логарифмов произведений размеров мультиплетов `CG и AT` на их количество `n` .
(8) Q1 = SUM(Ln(nCG*N))/ SUM(Ln(nAT*N))
Где : Q1 – `Первый` уровень эволюции .
nAT ,nCG - количество мультиплетов размером N для : - (nAT) и - nCG .
Так как этот критерий содержит в себе и `рудиментарные ` признаки – большие платформы внедрений , которые сильно влияют на уровень совершенства генома .
----
Так была закончена первая серия выдачи только что разработанной теории на форум для общего пользования.
Сама работа была уже прочитана несколькими большими учеными и доложена на семинаре в Болгарской Академии Наук. Отзывы были положительными,в смысле логичности предложенной модели. Но трудно воспринимаемой для того времени - "белковой парадигмы", в смысле восприятия.