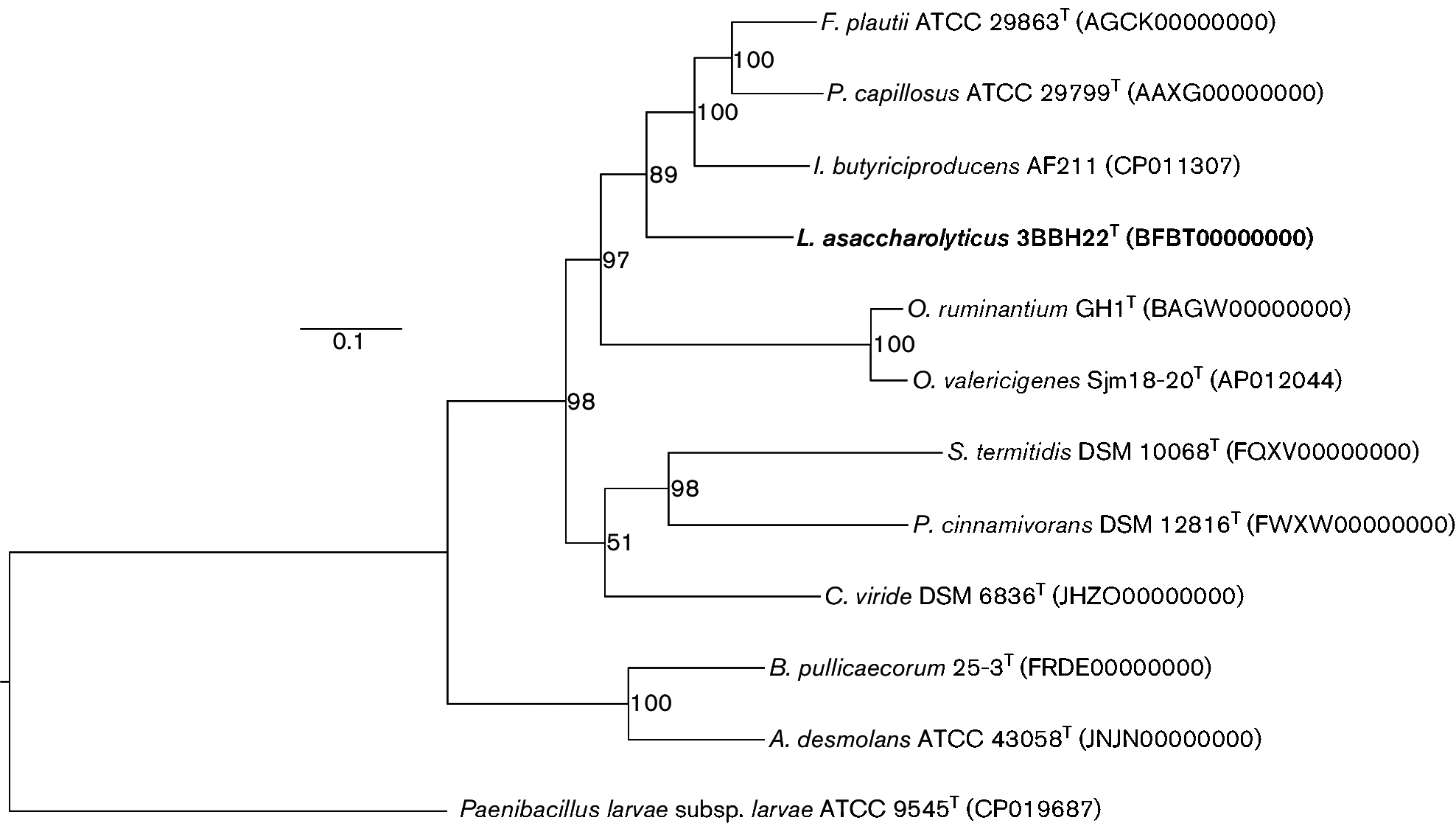
Облигатно анаэробная, грамположительная, неспорообразующая, прямая палочковидная бактерия, обозначенная штаммом 3BBH22 T , была выделена из фекального образца здоровой японской женщины. Анализ последовательности гена 16S рРНК показал, что штамм 3BBH22 T образовал монофилетический кластер с видами в родах Pseudoflavonifractor и Flavonifractor в семействе Ruminococcaceae и имел наибольшее сходство с Pseudoflavonifractor capillosus ATCC 29799 T (96,7 % сходства последовательности), за которым следует Flavonifractor plautii ATCC 29863 T (96,4 %). Ацетат и бутират были произведены штаммом 3BBH22 T в качестве конечных продуктов метаболизма. Основными клеточными жирными кислотами были C 14 : 0 , C 16 : 0 , C 18 : 1 ω9 c , C 16 : 0 диметилацеталь, C 18 : 0 и C 18 : 2 ω6,9 c . Дыхательных хинонов обнаружено не было. В отличие от F. plautii JCM 32125 T , штамм 3BBH22 T не разлагал кверцетин, один из флавоноидов. P. capillosus JCM 32126 T также не разлагал. Штамм 3BBH22 T отличался от P. capillosus JCM 32126 T неспособностью гидролизовать эскулин. Содержание G+C в геномной ДНК составляло 61,2 ± 1,0 моль%. На основании этих данных и филогенетического дерева, основанного на 89 белках, штамм 3BBH22 T представляет собой новый вид в новом роде семейства Ruminococcaceae , для которого предложено название Lawsonibacter asaccharolyticus gen. nov., sp. nov. Типовой штамм L. asaccharolyticus — 3BBH22 T (=JCM 32166 T =DSM 106493 T ).
В ходе наших попыток выделить микробы с полезными свойствами, такими как бактерии, продуцирующие бутират и разрушающие флавоноиды, мы успешно выделили пять новых видов: Butyricimonas faecihominis , Butyricimonas paravirosa , Parabacteroides faecis , Faecalimonas umbilicata и Parolsenella catena из человеческих фекалий. Помимо наших результатов, многочисленные новые виды были выделены из человеческого кишечника с помощью культуромики. Flavonifractor plautii , являющийся полифенол-превращающей бактерией, хорошо известен своей способностью метаболизировать кверцетин. Этот вид продуцирует бутират. F. plautii был предложен как единый вид Clostridium orbiscindens и Eubacterium plautii. Carlier et al. также предложили название Pseudoflavonifractor capillosus для обозначения неправильно классифицированного вида Bacteroides capillosus , который связан с F. plautii . Среди наших изолятов один штамм, обозначенный как штамм 3BBH22 T , был связан с F. plautii и P. capillosus филогенетически. В этом исследовании обсуждаются фенотипические, физиологические и филогеномные характеристики этого штамма.
Штамм 3BBH22 T был выделен из образца фекалий здоровой японки (41 год). Свежий образец фекалий был высеян на кровяной агар Brucella с гемином и менадионом (BB; среда JCM № 677). После 2–4 дней инкубации при 37 °C в газовой смеси H 2 /CO 2 /N 2 (1 : 1 : 8, по объему) штамм, обозначенный как 3BBH22 T , был выделен. Изолят рос анаэробно, но роста не наблюдалось после субкультивирования в микроаэробных условиях с использованием системы AnaeroPack-MicroAero (Mitsubishi Gas Chemical).
Хромосомную ДНК извлекали с помощью набора Genomic-tip 100/G (Qiagen). Для лизиса бактериальных клеток использовали лабиазу (5,0 мг мл −1 ; Cosmo Bio). Ген 16S рРНК анализировали, как описано ранее. Для штамма 3BBH22 T было определено около 1500 оснований последовательности гена 16S рРНК . Последовательности анализировали по базе данных EzTaxon ( www.ezbiocloud.net/identify ). Штамм 3BBH22 T показал наибольшее сходство с P. capillosus ATCC 29799 T (96,7 %), за ним следовали F. plautii ATCC 29863 T (96,4 %) и штамм Intestinimonas butyriciproducens SRB-521–5-I T (95,0 %). Связанные последовательности были выровнены с помощью программы clustal_x 2.0.12 и исправлены путем ручной проверки. Коэффициенты замены нуклеотидов ( значения K nuc ) были рассчитаны после устранения пробелов и неизвестных оснований. Филогенетические деревья были реконструированы методами объединения соседей и максимального правдоподобия (ML) с помощью программного обеспечения mega 7.0. Анализ Bootstrap был выполнен с 1000 повторными выборками для оценки достоверности топологий деревьев. Анализ последовательности гена 16S рРНК показал, что штамм 3BBH22 T образовал монофилетический кластер с P. capillosus ATCC 29799 T и F. plautii CCUG 28093 T ( рис. 1 ). Ген hsp60 оказался полезным филогенетическим маркером для грамположительных анаэробных бактерий, а также грамотрицательных анаэробных палочек. Частичный ген hsp60 был проанализирован, как описано ранее. Вкратце, частичный ген hsp60 был амплифицирован с помощью ПЦР с использованием ранее описанных праймеров H729 и H730 и секвенирован. Для анализа использовались нуклеотидные последовательности (552 п.н.) гена hsp60 из четырех штаммов. Последовательность гена hsp60 штамма 3BBH22 T показала 90,7, 88,7 и 85,4 % сходства с P. capillosus JCM 32126 T , F. plautii JCM 32125 T и I. butyriciproducens JCM 32124 T, соответственно. Результаты анализа последовательности гена hsp60 дополнительно подтверждают полезность гена hsp60 для классификации грамположительных анаэробных бактерий.

Весь геном штамма 3BBH22 T был секвенирован с использованием системы секвенирования PacBio RSII (Pacific Biosciences) от Takara. Чтения были собраны de novo с использованием Hierarchical Genome Assembly Process версии 3.0 ( hgap3.0 ) в smrt Analysis версии 2.3.0. Анализ последовательностей генома был выполнен с использованием Microbial Genome Annotation Pipeline (MiGAP; www.migap.org/index.php/en ). Нуклеотидные последовательности эталонных штаммов были получены из базы данных ncbi . Прогнозирование in silico значения гибридизации ДНК–ДНК (DDH) и попарной средней идентичности нуклеотидов (ANI), сравнивающее штамм 3BBH22 T с P. capillosus ATCC 29799 T , штаммом F. plautii YL31 и штаммом I. butyriciproducens AF211, соответственно, было выполнено с использованием калькулятора расстояний от генома к геному (GGDC) версии 2.1 (использованный инструмент локального выравнивания был blast +; https://ggdc.dsmz.de/ggdc.php #) и калькулятора ANI ( http://enve-omics.ce.gatech.edu/ani/ ), и полученные значения составили 22,6 % (с использованием формулы 2), 23,4 и 21,8 % соответственно для GGDC и 80,04, 82,08 и 79,51 %, соответственно, для ANI. Современный конвейер генов ядра бактерий ( ubcg ) ( www.ezbiocloud.net/tools/ubcg ) использовался для обнаружения 89 белков, что исключило три белка, LigA, PyrG и RplL из 92 белков набора данных ubcg , в геномах штамма 3BBH22 T и связанных таксонов. Неоднозначно выровненные позиции были устранены. Сцепленное дерево ML было реконструировано с помощью бутстреп-анализа (100 повторных выборок) с помощью RAxML . При анализе последовательности гена 16S рРНК таксономическое положение штамма 3BBH22 T не было ясным, тогда как филогенетическое дерево, основанное на 89 белках ( рис. 2 ), четко отделило штамм 3BBH22 T от других родственных видов ( F. plautii , I. butyriciproducens и P. capillosus ).
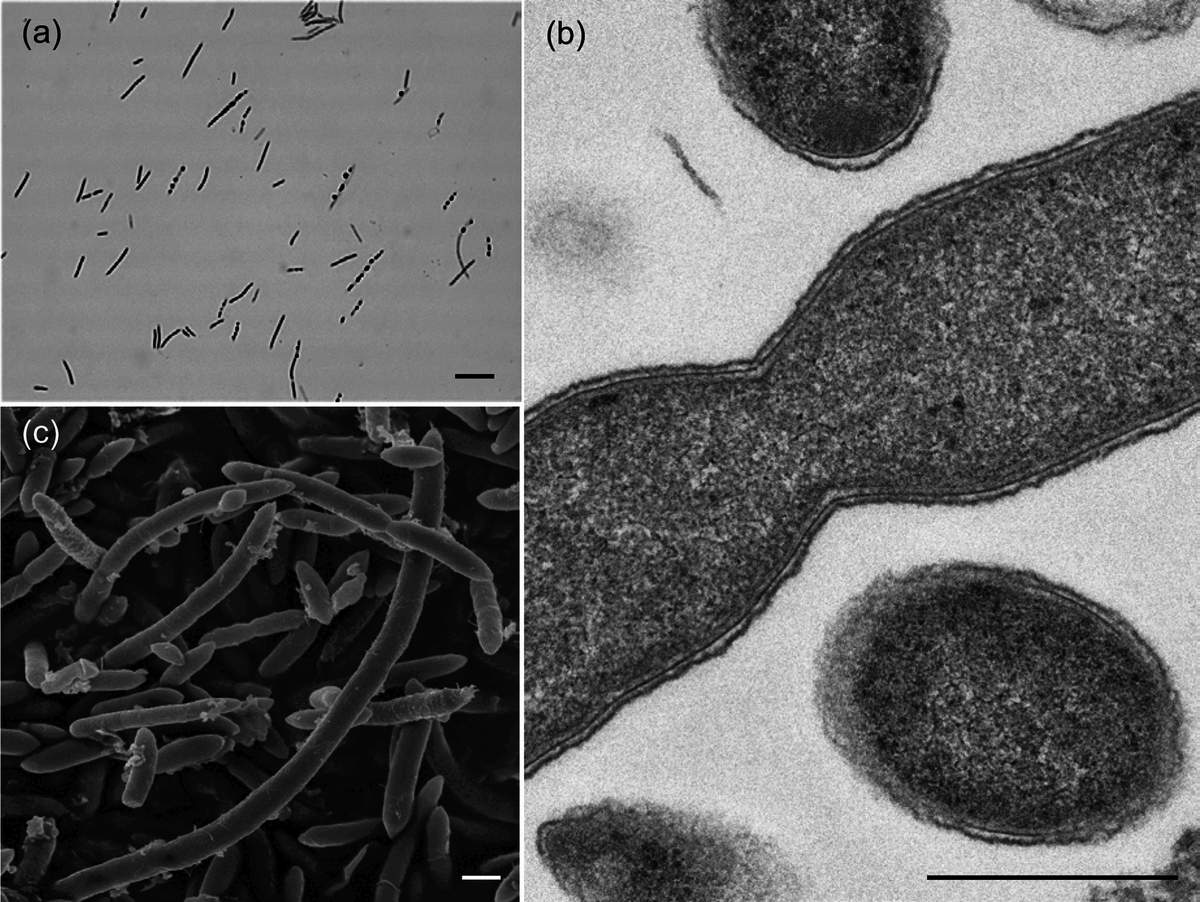
Штаммы, используемые в настоящем исследовании, выращивались на агаре BB в течение 2–4 дней при температуре 37 °C в газовой смеси H 2 /CO 2 /N 2 (1 : 1 : 8, по объему). В качестве контрольных штаммов использовались I. butyriciproducens JCM 32124 T , F. plautii JCM 32125 T и P. capillosus JCM 32126 T . Характеристика штамма 3BBH22 T и родственных видов проводилась в тех же условиях. Штаммы инкубировались при разных температурах (10, 20, 30, 37 и 45 °C) для изучения роста бактерий. Штамм 3BBH22 T рос при температурах от 30 до 45 °C с оптимальным ростом при 37 °C. Оптимальный pH (pH 4,0, 5,0, 6,0, 7,0 и 8,0) для роста определяли, как описано ранее. Штамм рос при pH 6,0–8,0. Оптимальным pH был pH 7,0. Клетки из культур, выращенных на агаре BB в течение 2 дней при 37 °C в анаэробных условиях, тестировали на окрашивание по Граму. Морфологию клеток после 2 дней культивирования на пластинах агара BB наблюдали с помощью фазово-контрастной микроскопии ( biophot ; Nikon), просвечивающей электронной микроскопии (ПЭМ; JEM-1400Plus; jeol ) и сканирующей электронной микроскопии (СЭМ; JSM-6340F; jeol ). Подготовка образцов для ТЭМ и СЭМ была описана ранее. Клетки штамма 3BBH22 T были облигатно анаэробными, непигментированными, прямыми палочками, отрицательно окрашенными по Граму. Клетки на агаре BB имели размер 0,8–1,5 (в среднем 1,0)×2,4–20 (в среднем 10,0) мкм и встречались поодиночке или парами с набуханием или без него ( рис. 3а ). Колонии на чашках с агаром BB после 4 дней инкубации при 37 °C в анаэробных условиях имели диаметр 0,5–1 мм, серые или серо-белые, круглые, слегка эрозированные, слегка заостренные и гладкие. Изображения ТЭМ и СЭМ клеток штамма 3BBH22 T показаны на рис. 3 (b, c). Типичные структуры спор, такие как описанные Брауном и др . , не наблюдались. В этом исследовании спорообразование не наблюдалось у F. plautii JCM 32125 T и P. capillosus JCM 32126 T с использованием световой микроскопии, как сообщают Карлье и др.. Хотя сообщалось, что небольшие терминальные и субтерминальные эндоспоры были обнаружены после 7 дней роста штамма I. butyriciproducens SRB-521-5-I T, спорообразование не наблюдалось у I. butyriciproducens JCM 32124 T . Карлье и др. исследовали наличие специфичного для споруляции гена spo0A для F. plautii DSM 4000 T и P. capillosus CCUG 15402 A T . Хотя споруляция не наблюдалась, эти два штамма обладали геном spo0A . Как и в случае с F. plautii и P. capillosus , ген spo0A был обнаружен в проекте генома штамма 3BBH22 T . Образование спор может быть вызвано некоторыми факторами окружающей среды. Структура клеточной стенки грамположительного типа была обнаружена в ТЭМ клеток штамма 3BBH22 T ( рис. 3b ), хотя его окраска по Граму была отрицательной. Это открытие похоже на открытие, полученное для F. plautii. Результаты этого исследования показывают, что штамм 3BBH22 T является неспорообразующей, грамположительной бактерией. Штамм 3BBH22 T был неподвижен, что было определено с помощью теста на подвижность, в котором пробирки, содержащие полутвердую анаэробную среду Гифу (0,15 или 0,5 % агара; Nissui), были инокулированы уколом и инкубированы при температуре 37 °C в течение 3 дней. Способность переносить желчь была проверена на агаре Bacteroides bile aesculin. На агаре Bacteroides bile aesculin рост I. butyriciproducens JCM 32124 T и F. plautii JCM 32125 T не был ингибирован в присутствии 0,5 % (w/v) Difco-Oxgall (BD), что эквивалентно 5 % (w/v) желчи для тестирования устойчивости к желчи. Эти два штамма слабо росли в присутствии 2 % (w/v) Oxgall. С другой стороны, рост штамма 3BBH22 T и P. capillosus JCM 32126 T был ингибирован в присутствии 0,5 или 2 % (w/v) Oxgall. Гидролиз эскулина был положительным только для P. capillosus JCM 32126 T .
Биохимические реакции определялись в двух экземплярах с использованием набора для анаэробного тестирования API 20A, системы API ZYM и набора для идентификации анаэробов Rapid ID 32A (bioMérieux) соответственно в соответствии с инструкциями производителя; результаты приведены в описании вида. Штамм 3BBH22 T оказался асахаролитическим при тестировании с использованием набора для анаэробного тестирования API 20A. При использовании бульона пептон-дрожжевой экстракт-глюкоза (PYG) (1 %, вес/объем каждого) глюкоза ферментировалась слабо. Штамм 3BBH22 T можно было отличить от P. capillosus JCM 32126 T по его неспособности гидролизовать эскулин ( таблица 1 ). При использовании системы API ZYM и набора для идентификации анаэробов Rapid ID 32A штамм 3BBH22 T был менее реактивным, чем контрольные штаммы. Тесты на чувствительность к противомикробным препаратам проводились с использованием различных противомикробных препаратов (BBL Sensi-Disc Susceptibility Test Discs; BD), включая амоксициллин, бацитрацин, хлорамфеникол, ципрофлоксацин, колистин, эритромицин, гентамицин, канамицин, неомицин, окситетрациклин, пенициллин, стрептомицин, сульфаметоксазол с триметопримом (SXT) и ванкомицин. Штамм 3BBH22 T был восприимчив к амоксициллину, бацитрацину, хлорамфениколу, эритромицину, окситетрациклину, пенициллину и ванкомицину. Штамм 3BBH22 T был устойчив к ципрофлоксацину, колистину, гентамицину, канамицину, неомицину, стрептомицину и SXT.
Органические кислоты конечных продуктов метаболизма в бульоне PYG анализировали, как описано Прамоно и др .. Выделение CO2 анализировали , как описано Сакамото и др . . Только F. plautii JCM 32125 T и P. capillosus JCM 32126 T продуцировали CO2 . Среди протестированных штаммов I. butyriciproducens JCM 32124 T , F. plautii JCM 32125 T и штамм 3BBH22 T продуцировали бутират ( таблица 1 ). Через 7 дней I. butyriciproducens JCM 32124 T продуцировал только 163,7 мМ бутирата. F. plautii JCM 32125 T продуцировал 23,1 мМ ацетата и 156,1 мМ бутирата. Штамм 3BBH22 T продуцировал 159,2 мМ ацетата и 30,1 мМ бутирата. Напротив, P. capillosus JCM 32126 T продуцировал 47,1 мМ ацетата и 14,7 мМ формиата. Кроме того, два штамма, F. plautii JCM 32125 T и P. capillosus JCM 32126 T , продуцировали незначительные количества лактата (1,9 и 3,6 мМ соответственно). Ген, кодирующий фермент, участвующий в производстве бутирата, бутираткиназу (EC 2.7.2.7), был обнаружен в проекте генома штамма 3BBH22 T . Были некоторые расхождения по сравнению с результатами, представленными в этом исследовании и предыдущих отчетах . Согласно предыдущим сообщениям, I. butyriciproducens продуцировал бутират и ацетат в бульоне Уилкинса-Чалгрена-Анаэробы, а P. capillosus продуцировал ацетат и сукцинат в бульоне TGY.
Метиловые эфиры жирных кислот (МЭЖК) были получены из примерно 40 мг влажных клеток, выращенных на агаре BB при 37 °C в течение 4 дней (все штаммы достигли стационарной фазы роста; кривые роста определялись путем подсчета колониеобразующих единиц), путем омыления, метилирования и экстракции с использованием незначительных модификаций метода Миллера. Профили жирных кислот клеток определялись с использованием версии 6.2B Системы идентификации микроорганизмов Sherlock ( midi ) и версии 3.80 базы данных bhibla . Основными клеточными жирными кислотами (>10 %) штамма 3BBH22 T были C 14 : 0 , C 16 : 0 , C 18 : 1 ω9 c , C 16 : 0 диметилацеталь (ДМА), C 18 : 0 и C 18 : 2 ω6,9 c ( таблица 2 ). В этом исследовании основными клеточными жирными кислотами P. capillosus JCM 32126 T были C 16 : 0 DMA, C 14 : 0 и C 18 : 1 ω9 c . Это был аналогичный результат для P. capillosus CCUG 15402 A T , о котором сообщали Kläring et al ., за исключением более высокой доли C 18 : 1 ω9 c . Доля C 16 : 0 , которая является одной из основных клеточных жирных кислот P. capillosus, о которой сообщают Карлье и др., была немного ниже (<10 %; 9,7 ± 0,7), чем пороговое значение основных компонентов, определенное в этом исследовании. В F. plautii JCM 32125 T основными клеточными жирными кислотами были C 16 : 0 , C 14 : 0 , C 16 : 0 DMA и C 18 : 0 DMA. Этот результат был почти таким же, как в предыдущих отчетах. Основные клеточные жирные кислоты I. butyriciproducens JCM 32124 T были аналогичны таковым штамма I. butyriciproducens SRB-521-5-I T, о котором сообщают Клеринг и др ., хотя были некоторые различия в пропорциях. Изопреноидные хиноны были извлечены и проанализированы, как описано Комагатой и Сузуки на наличие менахинонов и убихинонов. Ни менахиноны, ни убихиноны не были обнаружены ни в одном из протестированных штаммов.
Поскольку сообщалось о деградации кверцетина, одного из флавоноидов, F. plautii, мы проверили способность штамма 3BBH22 T деградировать кверцетин. Деградация кверцетина проверялась с помощью теста на гашение флуоресценции в соответствии с предыдущими исследованиями. Штамм 3BBH22 T не деградировал кверцетин. Среди протестированных штаммов только F. plautii JCM 32125 T был способен деградировать кверцетин, как сообщалось ранее. Эта особенность была описана как важная хемотаксономическая характеристика рода Flavonifractor.
Состав оснований ДНК для штамма 3BBH22 T и других референтных штаммов (за исключением F. plautii JCM 32125 T ) определяли методом высокоэффективной жидкостной хроматографии Тамаоки и Комагаты, используя смесь 0,02 M NH 4 H 2 PO 4 и ацетонитрила (20 : 1, об./об.) в качестве растворителя для элюирования. Содержание ДНК G+C штамма 3BBH22 T составило 61,2±1,0 мол.%. Это значение было почти таким же, как содержание ДНК G+C других штаммов ( I. butyriciproducens JCM 32124 T ; 61,5±0,2 моль%, F. plautii DSM 4000 T; 61,6 моль%, P. capillosus JCM 32126 T ; 61,0±0,4 моль%). Каждое значение представляет собой среднее значение ± sd трех экспериментов. Экспериментально определенное содержание ДНК G+C штамма 3BBH22 T (61,2±1,0 моль%) было немного выше, чем рассчитанное из черновой последовательности генома штамма 3BBH22 T (58,4 моль%) ( Таблица 1 ). Это может быть связано с неполнотой последовательности генома.
На основании физиологических и биохимических характеристик, а также филогенетического дерева, основанного на 89 белках, штамм 3BBH22 T представляет собой новый вид в новом роде семейства Ruminococcaceae , для которого предложено название Lawsonibacter asaccharolyticus gen. nov., sp. nov.
Описание Lawsonibacter gen. nov.
Lawsonibacter (Law.so.ni.bac′ter. NL муж. сущ. бактерия — палочка; NL муж. сущ. Lawsonibacter — палочка, названная в честь Пола А. Лоусона, Университет Оклахомы, за его большой вклад в таксономию и филогению микроорганизмов).
Грамположительные, облигатно анаэробные, неподвижные, неспорообразующие палочки. Асахариолитические; глюкоза может слабо ферментироваться. Филогенетически связаны с представителями родов Intestinimonas , Flavonifractor и Pseudoflavonifractor в семействе Ruminococcaceae . Типовой вид — Lawsonibacter asaccharolyticus .
Описание Lawsonibacter asaccharolyticus sp. nov.
Lawsonibacter asaccharolyticus [a.sac.cha.ro.ly'ti.cus. гр. прив. а , нет; гр. н. сакчар , сахар; НЛ маска. прил. lyticus (от греч. masc. прил. lytikos ), способный разрыхлять, способный растворяться; НЛ маска. прил. asaccharolyticus , не переваривающий сахар].
Клетки представляют собой непигментированные прямые палочки, встречающиеся поодиночке или парами с набуханием или без него, размером 0,8–1,5 (в среднем 1,0)×2,4–20,0 (в среднем 10,0) мкм. Споры не образуются, но присутствует специфичный для споруляции ген spo0A . Растет при температуре 30–45 °C (оптимум 37 °C) и pH 6,0–8,0 (оптимум pH 7,0). Колонии на чашках с агаром BB после 4 дней инкубации при температуре 37 °C в анаэробных условиях имеют диаметр 0,5–1 мм, серые или грязно-бело-серые, круглые, слегка эрозированные, слегка заостренные и гладкие. Роста не происходит на среде, содержащей 20 % (мас./об.) желчи. Сероводород, индол, каталаза и уреаза не продуцируются. Эскулин не гидролизуется. Желатин не переваривается. Не способен расщеплять кверцетин. Основные конечные продукты — ацетат и бутират. Преобладающие клеточные жирные кислоты — C 14 : 0 , C 16 : 0 , C 18 : 1 ω9 c , C 16 : 0 DMA, C 18 : 0 и C 18 : 2 ω6,9 c . Дыхательные хиноны не обнаружены. Асахариолитические при тестировании с использованием набора API 20A; глюкоза может слабо ферментироваться в бульоне PYG. Слабая активность получена с использованием набора Rapid ID32A для щелочной фосфатазы, аргинин ариламидазы, лейцин ариламидазы, тирозин ариламидазы и серин ариламидазы. Манноза и раффиноза не ферментируются. Отрицательные реакции получены для уреазы, аргининдигидролазы, α -галактозидазы, β -галактозидазы, 6-фосфо- β -галактозидазы, α -глюкозидазы, β-глюкозидазы , α -арабинозидазы, β -глюкуронидазы, N -ацетил- β -глюкозаминидазы, декарбоксилазы глутаминовой кислоты, α -фукозидазы, восстановления нитрата, продукции индола, пролинариламидазы, лейцилглицинариламидазы, фенилаланинариламидазы, пироглутаминовой кислоты ариламидазы, аланинариламидазы, глицинариламидазы, гистидинариламидазы и глутамилглутаминовой кислоты ариламидазы. Положительные реакции также получены с использованием системы API ZYM для эстеразы, эстеразолипазы, кислой фосфатазы и нафтол-AS-BI-фосфогидролазы. Отрицательные реакции получены для щелочной фосфатазы, липазы, лейцинариламидазы, валинариламидазы, цистинариламидазы, трипсина, химотрипсина, α - галактозидазы, β -галактозидазы, β -глюкуронидазы, α -глюкозидазы , β - глюкозидазы, N -ацетил- β -глюкозаминидазы, α -маннозидазы и α -фукозидазы. Организм чувствителен к амоксициллину, бацитрацину, хлорамфениколу, эритромицину, окситетрациклину, пенициллину и ванкомицину.
Типовой штамм 3BBH22 T (=JCM 32166 T =DSM 106493 T ) был выделен из человеческих фекалий. Содержание ДНК G+C в типовом штамме составляет 61,2±1,0 моль%.