Глава 18: Палеогеновый период.
Палеогеновый период (палеоген) — первый геологический период кайнозоя. Начался 66,0 млн. лет назад, закончился 23,03 млн. лет назад. Продолжался, таким образом, около 43 млн. лет. Именно в данный период происходит становление и рассвет млекопитающих, как главного наземного вида.
География.
На протяжении всего палеогенового периода Гондвана и Лавразия продолжали распадаться на составляющие. Северный суперконтинент Лавразия постепенно разделился на Северную Америку и Евразию. Гондвана окончательно распалась на Южную Америку, Африку, Австралию, Антарктиду и п-ов Индостан, который стремительно направлялся к южноазиатской части Евразии. За 40 млн. лет он благополучно достиг и воссоединился с ней. Жестокость столкновения привела к появлению самых высоких гор на Земле - Гималаев. Антарктида повернула к южному полюсу, а Южная Америка медленно шла на соединение с Северной.
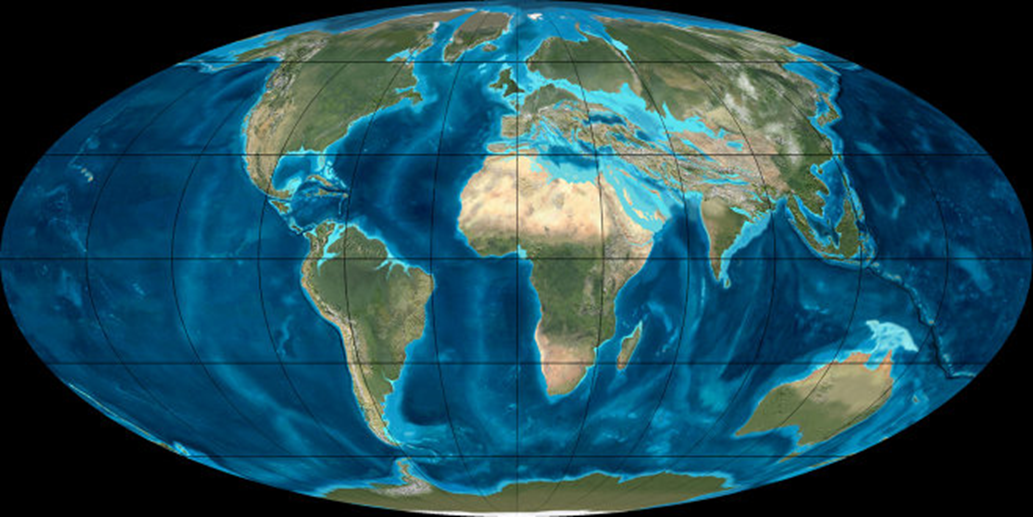
Гренландия окончательно отделились от континента Евразия. Тем самым закончилось формирование Атлантического океана на всем протяжении от Шпицбергена до Антарктиды. К окончанию палеогена ширина атлантического океана уже составляла от 1 тыс. до 2,5 тыс. км. Другие молодые океаны – Индийский и Северный Ледовитый -- продолжают расширяться и углубляться. Одновременно завершилось расширение Лабрадорского моря и моря Баффина между Северной Америкой и Гренландией. В Северной Америке вздымание Кордильер вызвало исчезновение остатков внутреннего морского бассейна. В Африке в палеогене существовали обширные внутренние моря делившие континент на несколько частей.
Климат.
В период палеогена воздушные массы, равно как и подводные течения, беспрепятственно и равномерно распределялись между всеми земными зонами, и поэтому климат был в основном теплым и влажным по всей поверхности. Но к концу периода средняя температура в пределах северных и южных широт упала настолько, что в зимние периоды на территории Антарктиды стали все чаще замерзать водоемы, а по всей площади Северного моря стали образовываться огромные блуждающие льдины, из которых впоследствии сформировались полюсные ледовые шапки.
Животный мир.
В морях и океанах на смену вымерших групп мезозойских животных пришли новые группы, получившие с началом палеогена огромный толчок в развитии. Особенно распространились фораминиферы-нуммулитиды, которые достигают нескольких сантиметров (в исключительных случаях — даже 19 см) в диаметре, что очень много для одноклеточных. Именно из известняков этих организмов были сложены древние египетские пирамиды. Наряду с ними были широко распространены и такие известковые одноклеточные водоросли, как кокколитофориды. Также значимая роль принадлежит радиоляриям, диатомным водорослям и микроскопическим кремниевым жгутиковым.
Дно океанов и морей покрылось новыми губками. Многочисленны были и кораллы, в основном относящиеся к склерактиниям. Имели место как мелководные, так и глубоководные их разновидности. Почти все рифовые коралловые массивы нынешнего тропического пояса зародились в середине палеогена. Из членистоногих известны остракоды. Меньшим распространением пользовались десятиногие ракообразные, в частности пресноводные и морские раки и креветки. Роль мшанок и брахиопод по сравнению с более ранними эпохами существенно сократилась. Из других беспозвоночных были широко распространены неправильные морские ежи.
Постепенно место вымерших аммонитов (ранее самой распространенной группы моллюсков) заняли новые виды брюхоногих и двустворчатых моллюсков. Больше всего было улиток. Они обитали не только в морях с нормальной соленостью, но и в солоноватых и пресных бассейнах. Некоторые моллюски освоили приморские низменности. Появились первые джулииды – пожалуй, самые интересные улитки в природе, они по виду и образу жизни ничем не отличаются от двустворчатых моллюсков, и даже их панцирь состоит из двух створок, увидев ее неспециалист и не поймет, что перед ним улитка, а не устрица. Значительным распространением пользовались и двустворчатые моллюски. Начинается расцвет головоногих моллюсков с редуцированной или вовсе исчезнувшей раковиной — осьминогов, каракатиц и кальмаров. Ранее известные головоногие моллюски балансировали на грани вымирания, известен лишь один род палеоценовых наутилоидов.
Исчезновение ихтиозавров, плезиозавров и других морских форм жизни, вымерших в конце мелового периода, оставило в пищевых цепях морей и океанов множество пробелов. Постепенно эти пробелы заполнились новыми группами рыб и акул. Среди палеоценовых рыб доминируют лучеперые рыбы, вымирание аммонитов и морских рептилий явно пришлось им по нраву. Среди них присутствуют уже почти современные окуни, селедки, сомы и щуки. Акулы палеогена становятся все более похожи на современных. Появились первые современные роды: леопардовая акула и ржавая акула-нянька. Самой большой палеогеновой акулой являлся отодус, достигавший в длину 9 м, тогда, как другие акулы были гораздо меньше – 1,5-3,2 м. Наряду с такими рыбами, как скаты и акулы, в океанических глубинах возникли и первые разновидности китовых, сирен и дельфиноподобных. Они стали первыми млекопитающими, решившими освоить океаническую и морскую водную среду. Есть утверждение, что первые предки китов произошли от парнокопытных.
Увеличивается разнообразие насекомых. В разы стало больше клещей, предков комаров и прочих кровососущих насекомых. С дальнейшим развитием цветковых растений все больше развивались и цветковые насекомые. Процветали бабочки и пластичатокрылые, жуки и всевозможные земляные виды членистоногих.
От земноводных после мелового катаклизма остались лишь лягушки, жабы и немногочисленные гигантские саламандры. Пережили меловое потрясение и некоторые разновидности рептилий, такие как черепахи, змеи, крокодилы и ящерицы. Крокодилы не только смогли выжить после мел-палеогенового вымирания, но фактически процветали. Размеры палеоценовых крокодилов составляли от 1 до 4,5 м, не все они были водными хищниками, некоторые бегали по земле, а pristichampsus умел даже бегать на двух ногах, как динозавр. Цератозух – единственный известный науке крокодил с рогами, зачем ему нужны были рога – науке неизвестно. Упомянем еще небольших водных рептилий хористодер (choristodera), похожих на крокодилов, но не родственных им. Чешуйчатые пресмыкающиеся породили самую большую змею за всю историю Земли – титанобоа длиной до 15 м, диаметром один метр в самом широком месте и весом до 1135 кг. Один из представителей чешуйчатых млекопитающих дожил до наших дней - двуходка rhineura.
Некоторые палеонтологи утверждают, что в первый миллион лет палеоцена кое-где еще встречались единичные виды динозавров, например, критозавр, чьи кости нашлись в палеоценовых отложениях. Одно из объяснений этой находки – кости динозавров занесло в более верхние слои осадочных пород каким-то геологическим процессом уже после окаменения. Но даже если динозавры реально жили в начале палеоцена, длилось это недолго.
После мел-палеогенового вымирания освободилось множество жизненных ниш, которые ранее были заняты динозаврами. С наименьшими потерями это массовое вымирание пережили теплокровные млекопитающие, которые и стали полноправными властителями земли. С этого периода начинается их бурный расцвет, который происходит и сегодня. В палеогене существовали как животные, которые впоследствии вымерли и теперь считаются ископаемыми, так и такие животные, которые стали предками многих современных животных. Например, в этот период появились предки носорогов, лошадей, свиней, верблюдов, кроликов, всех хищных, сумчатых, парнокопытных и непарнокопытных животных. Млекопитающие палеогенового периода подразделялись на яйцекладущих, сумчатых и плацентарных.
Первые яйцекладущие появились еще в середине мелового периода. Это наиболее примитивная группа млекопитающих. Они размножались, откладывая яйца; нередко их тело покрывала чешуя; строение черепа напоминало строение черепа пресмыкающихся. Со времен палеоцена до наших дней дожили лишь три их вида: два вида ехидн, или колючих муравьедов, и один вид утконосов. Все они встречаются только в Австралии и на Новой Гвинее.
Первые сумчатые обитали в Северной Америке с середины мелового периода. Позже они распространились по всем материкам, за исключением Африки и Азии. Сумчатые более высокоорганизованные животные, чем яйцекладущие, и в отличие от них имели постоянную температуру тела. Среди сумчатых млекопитающих были травоядные. Они напоминали современных кенгуру и сумчатых медведей. Были и хищники: сумчатый волк и сумчатый тигр. Некоторые сумчатые приспосабливались к жизни на деревьях. Они рождали недоразвитых детенышей, которых затем продолжительное время вынашивали в кожных сумках на животе. Многие из них питались лишь одним видом пищи, например, коала - только листьями эвкалиптов. Все это, наряду с другими примитивными чертами организации, привело к вымиранию сумчатых. Только в Австралии, рано отделившейся от других материков, эволюционный процесс как бы замер. Тут, царство сумчатых, сохранилось до наших дней.
В то время как яйцекладущие и сумчатые вырабатывали собственные способы размножения, еще одна группа млекопитающих (плацентарные) начала производить на свет потомство совсем иным путем. В основе их "метода" лежал особый орган — так называемая плацента. Главная же "идея" состояла в том, чтобы детеныш оставался внутри материнского организма, пока не достигнет сравнительно высокого уровня развития. У этого нового способа размножения были вполне очевидные преимущества. Рождаясь куда более "созревшими" детеныши плацентарных млекопитающих имели гораздо лучшие шансы выжить. Кроме того, плацентарные значительно усовершенствовали методику выкармливания своих новорожденных отпрысков и выработали новый тип поведения, рассчитанный на длительные периоды ухода за молодняком.
В палеоцене многие плацентарные млекопитающие оставались маленькими животными, чем-то похожими на своих меловых предков. Однако они очень быстро эволюционировали, и вскоре начали составлять серьезную конкуренцию сумчатым. Способность поддерживать постоянную температуру тела, прогрессивный способ размножения и крупный головной мозг позволили им стать процветающей группой животных и постепенно установить свое господство на всей поверхности земного шара. Ближе к середине палеогена начало происходить ярко выраженное разделение основной массы плацентарных млекопитающих на такие систематизированные группы, как хищные, хоботные, копытные, насекомоядные, приматоподобные, грызуны и пр. Но ввиду того, что в большинстве своем они не спешили с развитием то так и остались довольно примитивными, многие из них вымрут уже в будущем неогене.

Кондилартры – видимо, именно они были предками парнокопытных (к ним относят свиней, оленей, антилоп и др.) и непарнокопытных (лошади, тапиры, носороги), хоботных и даже наземных предков китов и сирен. Некоторые кондилартры преобразовали когти на пальцах в копыта, некоторые по-прежнему жили с когтями.
Самый большой надотряд плацентовых – лавразиотерии являвшиеся потомками мезозойских млекопитающих, обитавших в Евразии и Северной Америке. Начнем рассматривать основные отряды лавразиотериев с cimolesta. Этот отряд возник во второй половине мелового периода и вымер в середине кайнозоя. Животные были очень разнообразными, их размеры варьировались от 12 см до 2,5 м, вес – от 60 г до 650 кг. Мелкие симолесты были насекомоядными, крупные – растительноядными, несмотря на то, что имели клыки и когти. Дело в том, что в эволюционной гонке палеоцена побеждали не те животные, которые лучше приспособились к своим экологическим нишам, а те, кто первыми успели эти ниши занять. Копытные хищники, а так же когтистые и клыкастые козлы являлись обычным делом для палеогена.
Второе место по численности родов занимал отряд мезонихий, это предки современных парнокопытных, а также китов. Синоникс входящий в этот отряд и имевший много мелких признаков, присутствующих у китообразных, и не присутствующих больше ни у кого считается непосредственным предком китов. Несмотря на копыта на лапах, первые мезонихии были хищниками, самые крупные достигали 1,5 м в длину. Самые первые примитивные млекопитающие-хищники — миациды, которые стали прародителями всех современных хищников. Судя по находкам, первые миациды были чем-то похожи на куницу и охотились на мелких животных и насекомых. В конце периода появляются более крупные хищники — энтелодоны и гиенодоны.
Гиенодоны
Основными представителями крупных растительноядных были непарнокопытные: тапирообразные, носорогообразные, бронтотерии, халикотерии, архаичные лошадиные. Неуклюжие травоядные млекопитающие, амблиподы, поедали листья и прочую растительность. Бивни и странные рога некоторых из них служили для защиты от хищников. Бронтотерии были широко распространены в середине палеогена по всем материкам и географическим зонам. В основном они питались сочной болотной растительностью и подолгу могли оставаться в воде. Также влажные и заболоченные места были полны эптелодонтами, аминодонтами и антракотериями (свинообразными животными, отдаленно напоминавшими гиппопотамов). В тропических лесах развились гигантские млекопитающие носороги индрикотерии, самые крупные сухопутные животные в те времена, которые достигали пяти метров в высоту и 20-ти тонн веса. Наряду с ними редколесья обживали и пресноводные черепахи. Начинали появляться такие новые формы, как первые виды слонов.
Индрикотерии
Южноамериканские копытные, в полном соответствии со своим названием, жили в Южной Америке. В начале кайнозойской эры она была изолирована от других континентов, и так получилось, что среди местных млекопитающих не нашлось достаточно продвинутых, чтобы быстро занять освободившиеся экологические ниши. Местные землеройки росли и специализировались в козлов, коров и носорогов, это был долгий и мучительный процесс. Эти звери были невелики (до 1,5 м в длину), у некоторых были клыки, как у современных кабанов. В конце концов, все они вымерли, когда Южная Америка соединилась с Северной и южноамериканским копытным пришлось конкурировать с «настоящими» копытными.
Вторым большим надотрядом палеогеновых плацентарных были Euarchontoglires – это общее название для грызунов, приматов и шерстокрылов. Они были маленькими (вес до 2,1 кг) и всеядными, многие жили на деревьях. У некоторых появился противопоставленный большой палец, эти существа разделились на два отряда, один из которых – приматы. Другими представителями палеоценовых лавразиотериев представленных по одному роду, являлись диноцераты – большие носорогоподобные травоядные, креодонты – альтернативный отряд специализированных хищников и панголины.
Но как бы не развивались все виды млекопитающих в палеогене птицы, а не млекопитающие, были доминирующими наземными животными на Земле (что не должно удивлять, учитывая, что они эволюционировали от недавно исчезнувших динозавров). Это были огромные нелетающие (бегающие) хищные птицы,которые внешне напоминали хищных динозавров, но полностью вымершие в том же палеогене,- диатрима, гасторнис и фороракос. Диатрима была 2 м в высоту с длинным, до 50 см клювом. На сильных лапах у нее имелось по четыре пальца с длинными когтями. Жила диатрима в засушливых степях, питалась мелкими млекопитающими и пресмыкающимися. Фороракос достигал 1,5 м в высоту. Его острый крючковатый полуметровый клюв был весьма грозным оружием. Длинные, сильные ноги фороракосов свидетельствуют о том, что они были прекрасными бегунами. По мнению некоторых исследователей, родиной этих огромных птиц была Антарктида, покрытая в то время лесами и степями. В джунглях Европы и Северной Америки охотился свирепый хищник гасторнис, достигавший высоты 2,2 метра.
Однако появились более разнообразные летающие веерохвостые птицы, которые во многом были похожи на современных птиц. Сюда входят гуси, утки, совы, фламинго, пеликаны, бакланы, буревестники, ржанки, казуары и пелагорнитиды – типа альбатросов, но очень большие и с зубами в клюве, а так же пресбиорнис – нечто вроде журавля, построенного на генетической базе гуся, вайману являвшимся предком всех пингвинов и страуса эму, этот род возник как раз в палеогене.
Растительный мир.
С наступлением палеогена опустевшая планета начинает медленно восстанавливаться от последствий катастрофы. В этом периоде подавляющее количество растительных форм стало покрытосеменными. Увеличивается разнообразие цветковых растений.
В зоне экватора на территориях материков произрастали влаголюбивые леса дождевой и муссонной направленности, во многом схожие с современными центральноамериканскими, западноафриканскими и индийскими джунглями. Основу данных лесов составляли пальмовые, фикусовые и сандаловые древесные разновидности. Далее к центральным областям эти леса сменяли саванны и редколесья. На территориях аридных поясов росли растения, характерные для саванн, в основном это были ксерофиты. На рубежах аридных поясов климат становился влажнее, что способствовало разрастанию растительности смешанных типов. По берегам водоемов раскинулись чащи дубовых узколистных лесов, под сенью которых произрастали лавровые. Кое-где такие леса часто перемежались с миртовыми и хвойными.
Ближе к полюсам в лесах начинали преобладать вечнозеленые дубы, миртовые, каштановые и кипарисовые деревья. Преобладали в них также секвойи, туя, араукарии, различные таксодиевые и лавровые. В некоторых местах еще сохранялись отдельные заросли цикадофитовых и гинкговых. В нижних частях нынешнего умеренного пояса попадались примеси тропических растений, но в основном до самых окраин Северного океана и на всей территории Антарктиды произрастали хвойно-широколиственные листопадные леса. Лишь в местах, где климат был чрезвычайно влажен, а дожди шли слишком часто хвойно-широколиственные листопадные леса сменялись зарослями секвой, таксодиума, тиса, сосновыми, кедровыми и пихтовыми лесами, каштановыми, кленовыми, платановыми и всевозможными ореховыми рощами. Но нередко между ними можно было повстречать и вкрапления пальмовых, миртовых, лавровых и магнолиевых.
Установившийся во времена палеогена теплый климат вполне способствовал процветанию лесов на всей территории Антарктиды. Ниже северного и выше южного полярных кругов резкий контраст между временами года вовсе отсутствовал, и поскольку на то время минусовых температур не существовало, это способствовало распространению лесных массивов по всем континентальным территориям. Листопадные леса заполярных районов сбрасывали листву не с наступлением холодов, поскольку температурный режим зимних периодов в ту пору был выше нулевой отметки, а с наступлением полярной ночи. Вегетация на этом этапе прекращалась и голые растения, пережив долгую ночь, вновь начинали расцветать и обзаводиться листвой лишь с первым полярным рассветом.
К концу палеогена развилось повсеместное похолодание, что обусловило значительное смещение климатических зон ближе к экватору. С наступлением холодов усилилось влияние хвойных, возникли новые разновидности покрытосеменных. Огромные массивы крайне южных и северных широт заросли березовыми, тополиными, ивовыми и ольховыми лесами. А в связи с расширением областей аридизации климата впервые к концу палеогена в зонах нынешнего умеренного климата возникли лесостепи и савано-степи. Появились первые травы, которые оказали значительное влияние на жизнь последующих периодов.