Читаем учебник “Молекулярная биология” Коничев А.С. Севастьянова Г.А. Глава 7. Репликация ДНК. (Стр. 207).
Основной синтез новых ветвей ДНК ведет ДНК-полимераза, но оказывается, что
“ДНК-полимеразы не способны инициировать синтез новых цепей ДНК, они могут лишь добавлять дезоксирибонуклеотидные звенья к З'-концу уже имеющейся полинуклеотидной цепи. Чтобы молекулы ДНК-полимераз могли начать синтез ДНК, им необходима затравка, или праймер (от англ. priтеr – затравка), короткий олигодезоксирибонуклеотид или олигорибонуклеотид, комплементарный соответствующему участку ДНК-матрицы, у которого на конце имеется свободная З'-ОН-группа”. (Стр. 209).
Как понимать: в природе существует и олигодезоксирибонуклеотид и олигорибонуклеотид для синтеза одной и той же ДНК? Например, один праймер участвует в синтезе материнской молекулы, а другой праймер работает в дочерной молекуле. Или в репликации одной ДНК участвует праймер одного и того же типа? Тогда по какому принципу выбирается тот или иной праймер?
Затравки в клетке не с неба сваливаются – их синтезирует фермент ДНК-праймаза.
“На стадии инициации репликации короткую РНК-затравку из рибонуклеозидтрифосфатов синтезирует фермент, называемый ДНК-праймазой. ДНК-праймаза может быть отдельным ферментом (как у бактерий) или входить в качестве субъединицы в ДНК-полимеразу (как у ДНК-полимеразы эукариот)”. (Стр. 209).
Видите, ДНК-праймаза синтезирует не ДНК-затравку, а РНК-затравку. То есть начальный участок реплицируемой ДНК не реплицируется в понимании данного слова, а транскрибируется, так как в затравке находится сахар не дезоксирибоза, а именно рибоза. В ДНК ее не должно существовать и поэтому этот праймер надо убрать. И как пишется в учебнике:
“В дальнейшем короткие праймеры замещаются сегментами ДНК”. (Стр. 209).
Чтобы заместить, сначала надо удалить эти рибонуклеозидтрифосфаты. Чем? Снова надо привлекать механизм коррекции? Но механизм коррекции удаляет один нуклеотид, а РНК-затравка хоть и короткая, но все же не один нуклеотид. Затем на освободившиеся основания матрицы надо посадить требуемые нуклеотиды, а это может делать только ДНК-полимераза. Что ее надо возвращать в начало строящейся ветви, пусть она достроит начало новой ветви? Или как закончить строительство новой ДНК? Какая-то путаница.
Но еще большие сложности возникают при синтезе молекулы на другой цепи.
“Поскольку две цепи в молекуле ДНК антипараллельны, а ДНК-полимеразы способны наращивать полинуклеотидную цепь только в направлении 5'->3', один и тот же фермент, ДНК-полимераза, не может обеспечить сборку дочерних цепей одновременно в направлениях 5'->3' и 3'—>5' в соответствии с перемещением репликативной вилки при расплетании биспиральной молекулы ДНК”. (Стр. 209).
Видите, какая возникает проблема: ДНК-полимераза может наращивать нуклеотиды только в одном направлении – от 5'->3', как показано на рисунке. В обратном направлении она эту операцию совершать не может. Выход из этой ситуации предложил ученый Оказаки.
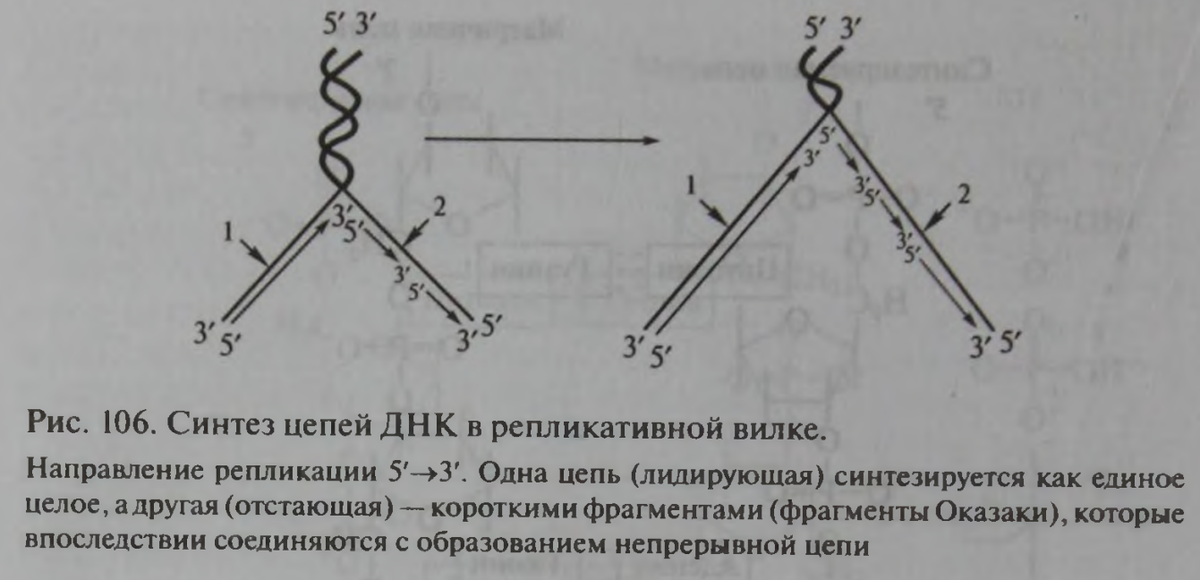
Он объяснил, что на лидирующей цепи наращивание нуклеотидов происходит непрерывно и при том чрезвычайно быстро. Как быстро, естественно, не объясняется, но из статьи “Полчаса митоза” видно, что процесс репликации длится десятки минут.
На отстающей цепи ситуация другая. ДНК-полимераза выжидает время пока ДНК не расплетется на 1000-2000 нуклеотидов. Затем ДНК-полимераза находит конец 5' и начинает с него синтез строящейся цепи в направлении к концу 3'. То есть синтез осуществляется фрагментами, которые затем сшиваются в целую ветвь, соединенную с отстающей ветвью, чем и образуют полноценную дочернюю ДНК.
К данной модели есть вопросы. Включается в работу при синтезе фрагмента Оказаки ДНК-праймаза? Или она в данном случае не работает? В клетках эукариот ДНК-праймаза в виде субчастицы, по мнению авторов книги, включена в состав ДНК-полимеразы.
Если ДНК-полимераза будет принимать часть цепи со стороны репликативной вилки, как затравку, то строить праймер не надо и ДНК-праймаза в данном случае не работает. Не будет ли возникать конфликт между этими двумя объектами? Один будет пытаться строить праймер, а другой, на этих же элементах, будет строить нормальную цепь.
Как-то не совсем понятно: обе цепи строит одна и та же ДНК- полимераза или это делает один тип полимераз? Может быть одна и та же ДНК-полимераза строит какой-то участок лидирующей цепи пока догонит репликативную вилку. Затем перемещается на отстающую цепь, достраивает ее до предыдущего фрагмента, уже построенного в предыдущем цикле и затем эта же ДНК-полимераза возвращается на лидирующую цепь и цикл повторяется.
Не угадал я, оказывается:
“Вновь образованная цепь, которая синтезируется непрерывно, получила название ведущей, или лидирующей, а другая, собираемая из фрагментов Оказаки, ведомой, или отстающей. Синтез каждого из этих фрагментов начинается с РНК-затравки. Такой механизм репликации называется прерывистым (или полунепрерывным). Ведущая цепь нуждается только в одном акте инициации, а для синтеза отстающей цепи должно произойти несколько актов инициации. Через некоторое время РНК-затравки (праймеры) удаляются, бреши «застраиваются» ДНК-полимеразой и фрагменты сшиваются специальным ферментом ДНК-лигазой в одну непрерывную отстающую цепь”. (Стр. 219).
Как же получается? Цепь разрывается и в разрыв помещается праймер или часть цепи перед фрагментом затирается, то есть с сахаро-фосфатного остова убираются азотистые основания, и на нем строится праймер? Второе невозможно, так как оно приводит к потере информации.
Я пытаюсь представить работу ДНК-праймазы и ДНК-полимеразы и вот что у меня получается. Само собой, понятно, что я могу ошибаться и представлять все не так, как представляют это ученые. Перед вами рисунок, на котором изображен кусок матрицы, равный 4 нуклеотидным парам. Во всей молекуле таких пар миллиарды или 2000-3000 в фрагментах Оказаки.
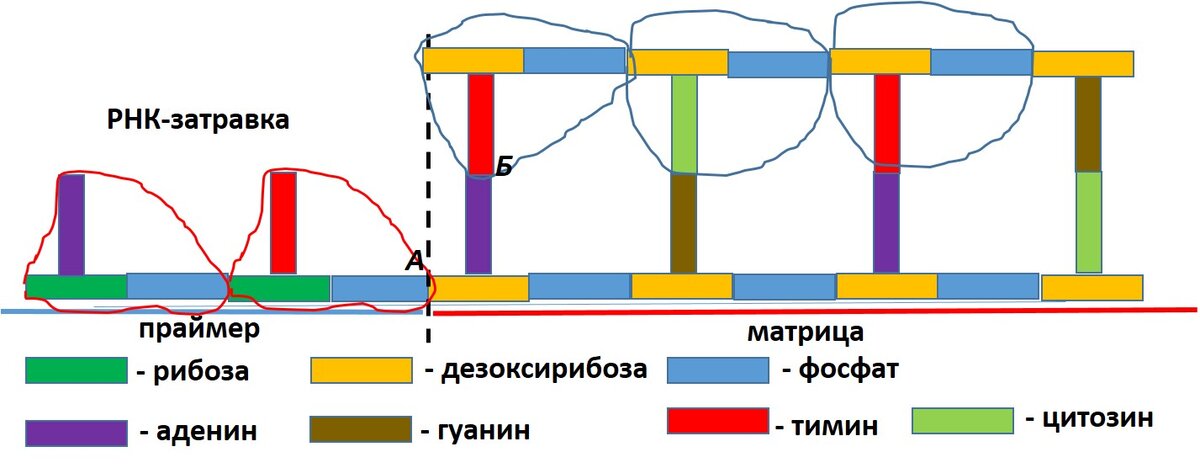
Задача ДНК-полимеразы состоит в том, чтобы на матрицу посадить комплементарные нуклеотиды, которые я обвел синей линией. Но она не может начать выполнять данную работу, пока не будет построен праймер в виде РНК-заправки. Праймер тоже состоит из нуклеотидов (я обвел их красной линией). Я нарисовал всего два нуклеотида, но в самом деле их больше по предположению ученых.
А теперь давайте хорошенько подумаем. О том, как появилась матрица мы поговорим ниже. Сейчас будем считать, что она плавает в растворе внутри клетки. Можно даже думать, что ее место примерно в том районе, где было ядрышко, в котором она жила в виде хромосом.
А где находилась в это время ДНК-полимераза? В начале матрицы или перед фрагментом Оказаки? Может быть, но вероятность этого мала. Предположим какие-то силы приблизили ДНК-полимеразу к началу матрицы или она сама приплыла. Есть ли такие силы, которые могли бы передвигать полимеразу, ученые не знают. В принципе они есть, но о ней ученый люд и не догадывается и в данном случае она не нужна.
Хорошо, допустим ДНК-полимераза оказалась в нужном месте. Теперь что? Субъединица , представляющая фермент ДНК-праймазу, должна из клеточного бульона отловить определенный или любой нуклеотид для РНК-затравки и соединить его с сахаром матрицы. Затем проделать такую же операцию с еще некоторым количеством нуклеотидов.
Да никакое животное не выложит в таком порядке синие и зеленые прямоугольники, кроме человека и то не каждого. А говорить о маленьком кусочке мяса и вовсе не следует.
После создания праймера ДНК-полимераза возвращается в начало матрицы или фрагмента и начинает добавлять на них комплементарные элементы. Вот что мешало полимеразе выполнять эту работу без праймера? Почему именно он инициировал работу полимеразы? Несомненно, что соединение в точке А может изменит состояние в точке Б. Нуклеотид – это молекула, а молекула, примерно, как паутина или сеть: потянешь в одном месте – произойдут некие изменения в другом месте. А есть ли такие размышления у авторов учебника, я не знаю.
В общем эта суета ДНК-полимеразы в режиме репликации кажется не правдоподобной. То есть такого движущегося объекте в клетке не существует. Да и согласно квантовым воззрениям такой объект и не нужен. Наращивание нуклеотидов осуществляется совершенно по другому принципу. Все освободившиеся концы азотистых оснований в результате расплетения под воздействием определенного излучения (потока энергии в виде фотонов различной энергии) светятся своим конкретным светом. То есть на них падают несколько видов энергии, а излучают они суммарную или разностную величину падающей энергии. Аденин светится одним цветом, тимин – другим и т. д. Цвет аденина поглощается только нуклеотидом с тимином и этот нуклеотид под гравитационным воздействием движется к светящемуся концу аденина, организует с ним связь, чем и прекращает свечение аденина.
Правда здесь существует вопрос: в каком порядке синтезируются нуклеотиды? Возможно они синтезируются отдельно и заблаговременно в клетке, а возможно, что к матрице сразу присоединяется один комплементарный элемент, затем к нему присоединяется дезоксирибоза и затем присоединяется фосфат. В любом случае происходит под воздействием фотонов. В том числе также реагируют и концы 3' и 5'.
Может ли участвовать в этом процессе ДНК-праймаза и ДНК-полимераза? Да, но только именно как фермент, то есть излучать тот или иной свет (поток энергии в виде фотонов). Об этом можно прочесть в статье о репликации.
У меня другие понятия о ферментах. Я больше склоняюсь к тому, что ферментами являются не полимеразы, праймазы, лигазы и тому подобное, а гистоны. Почему я так считаю? Потому что они есть в половых клетках и, соответственно, в зиготе и затем в дальнейших стволовых клетках. Все эти клетки успешно делятся. А всевозможных рибосом и –аз в зиготе или моруле нет. Гистонов несколько типов и из них можно получать богатый спектр излучений. Так это или не так, пока не известно.