«не обращай внимания на все оскорбления! ты прекрасна! у тебя даже хромосом больше, чем у тех, кто тебя оскорбляет!» (с) бессмертное с башорга
Известно, что все эукариоты имеют диплоидный набор хромосом. Ну, кроме простейших… И некоторых растений… И грибов… Членистоногих? Ну да, куча эукариот живет либо со случайным количеством хромосом, как например амёбы, либо имеют полиплоидию, как достаточно большое количество растений и грибов, либо вообще имеют кастрированный в результате «диминуции» геном, как рачки циклопы или аскариды. Но это все верно исключительно для соматических клеток (в случае многоклеточных организмов) либо для макронуклеуса (в случае одноклеточных простейших). В целом же, половой процесс, который таки показан для подавляющего большинства эукариот (кроме нескольких представителей протист, которых реально можно пересчитать по пальцам не очень внимательного столяра), требует диплоидного набора хромосом.
Что такое диплоидный набор хромосом? Это два гаплоидных набора хромосом. В случае высших эукариот, представителей которых можно разделить по их половой принадлежности, гаплоидный набор — это тот набор хромосом, который достается от одного из родителей. Т.е. два родителя (пока это еще так, как дальше пойдет эволюция — знают только власти ЕС) — два гаплоидных набора от каждого. И все бы ничего, но есть один нюанс. У многих видов половая принадлежность определяется комбинацией так называемых «половых» хромосом. У млекопитающих это всегда X и Y. Два икса — девочка, икс + игрек — мальчик.
И тут надо сделать лирическое отступление. Все мы… Ну ладно, может не все, но подавляющее большинство знает шутки про «лишнюю» хромосому. Надо заметить, что лишняя хромосома появляется в случае, когда при созревании гамет (яйцеклетки или сперматозоиды) случайно происходит нерасхождение хроматид. В результате, один из участников акта Яйцеклетка+Сперматозоид имеет эту самую лишнюю. Все знают синдром Дауна — это трисомия по 21 хромосоме. Есть еще синдром Эдвардса (18 хромосома) и Патау (13 хромосома). Для остальных хромосом синдромы не описаны, ибо трисомии по остальным хромосомам в подавляющем большинстве случаев летальны и развитие эмбриона прекращается еще на ранних стадиях. Это связано с таким понятием, как «доза гена». Как известно, хромосомы — это носители нашей генетической информации, т.е. всей совокупности генов. Каждый ген представлен определенным количеством копий: от 2-х (различные гены, отвечающие за регуляцию клеточного цикла, дифференцировку и т. д.) до нескольких тысяч (гены «домашнего хозяйства», т. е. участвующие в постоянной жизнедеятельности клетки, например в гликолизе или синтезе белков). Каждый ген характеризуется определенным уровнем «экспрессии», т. е. уровнем продукции кодируемого геном продукта (белка или какой-либо функциональной РНК). Избыток такого продукта может нарушить функционирование клетки самым драматическим образом, по-этому в разных типах клеток разные участки хромосом «запаковываются» в гетерохроматин*, чтобы блокировать доступ к определенным генам.
Теперь вернемся к нашим мальчикам и девочкам. В случае аутосом (все хромосомы, кроме X и Y) количество локализованных на них генов одинаково и у мальчиков, и у девочек. А вот в случае с Х у девочек всего в 2 раза больше, чем у мальчиков (Y в данном случае не рассматривается, ибо функциональных генов там примерно 5, и все они связаны исключительно с необходимостью запустить развитие эмбриона по мужскому пути). Как результат, в процессе эволюции выработался механизм инактивации «лишней» Х-хромосомы у женщин. Этот процесс связан с упаковкой одной из Х-хромосом в сверхплотный гетерохромаин, в результате которой достаточно большая хромосома превращается в тельце Барра — микроскопический «клубок» едва заметный в мискроскоп. Инактивация происходит на самых ранних этапах эмбрионального развития — либо сразу после оплодотворения яйцеклетки, либо после первого дробления, и у этого процесса есть одна особенность: упаковывается всегда одна и та же хромосома. Какая именно Х-хромосома — доставшаяся от отца или от матери — будет инактивирована, определяет случай, но в дальнейшем «потомство» именно этой хромосомы будет инактивироваться во всех клетках-потомках. Если инактивация произошла сразу после оплодотворения, то ВСЕ клетки организма имеют одну и ту же «рабочую» Х-хромосому и абсолютно неотличимы друг от друга по этому признаку. Однако в некоторых случаях такая инактивация происходит либо после первого дробления оплодотворенной яйцеклетки, либо после второго (т. е. на стадии 2-х или 3-х бластомеров**), и в данном случае в разных клетках могут инактивироваться разные Х-хромосомы. В результате получается мозаичный организм, где часть клеток имеет рабочую Х-хромосому от матери, а другая часть — от отца. Типичный пример такой мозаичности — черепаховые (трехцветные) кошки. Особенность черепаховых кошек в том, что не бывает 2-х кошек с одинаковым рисунком вследствие случайности распределения клеток, опредеяющих тот или иной цвет.
Ну и немного о механизмах, обеспечивающих избирательность инактивации Х-хромосомы. Одним из способов регуляции экспрессии генов является метилирование, т. е. модификация Цитозина путем присоединения метильной группы (СН3-).
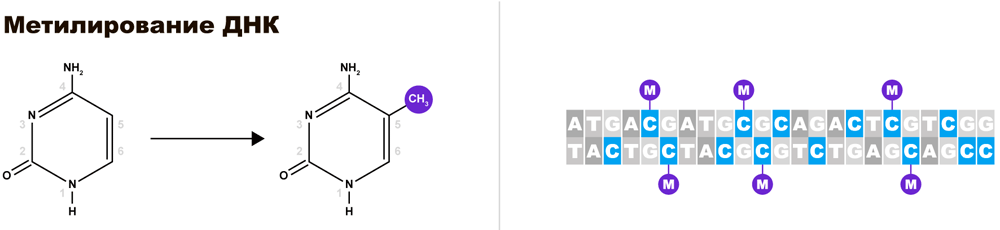
Избыточное метилирование является сигналом для упаковки метилированного участка хромосомы в гетерохроматин. При метилировании всей хромосомы — упаковывается вся хромосома. Именно так происходит первичная инактивация одной из Х-хромосом.
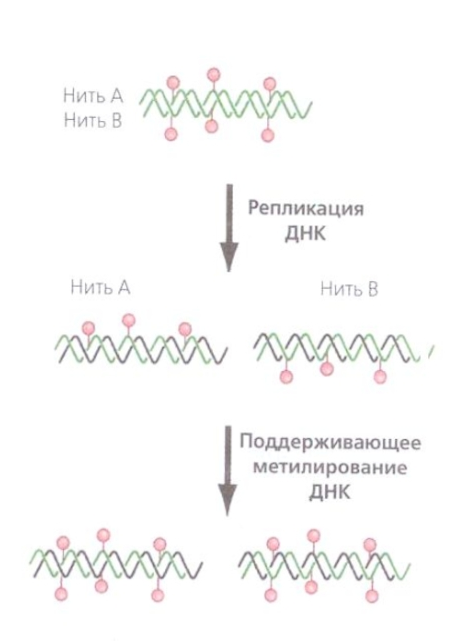
Как известно из школьного курса Биологии (в мое время это была биология 9-10 класс, как сейчас — ХЗ, ибо дети пока в 5-м, и этого еще не было) ДНК состоит из 2-х цепей. Метилируются обе цепи (как следует из рисунка). После удвоения (репликации) получается две двойных молекулы ДНК, в которых одна цепь метилирована, а другая, новосинтезированная, — нет. Но наличие метилированных оснований в одной из цепей является сигналом для метилирования этих же участков во второй. Так происходит «наследование» паттерна (pattern — шаблон, образец, рисунок) метилирования. Ну и, разумеется, если метилирована вся ДНК хромосомы, то получившаяся после репликации ДНК дочерних хромомосом тоже будет метилирована по всей длине и, как результат, эти дочерние хромосомы будут аккуратно сложены с то самое тельце Барра.
---------------------------------------------------------------------------------------------
*Гетерохроматин — очень плотно упакованный хроматин, в котором ДНК так облеплена белками и «свернута» в колечки, что доступ к ней становится невозможным, в результате все гены, локализованные в данном участке перестают работать.
**Бластомер — клетка, формирующая эмбрион на стадии «бластулы», т. е. до начала формирования специализированных тканей. Иными словами, все бластомеры одинаковы и неотличимы друг от друга, и, теоретически, каждый бластомер, будучи изъят из формирующегося эмбриона, может развиться в полноценный организм. Собственно, так и получаются близнецы — бластула на стадии двух или трех бластомеров разваливается, и каждый отвалившийся бластомер дает отдельный эмбрион.